ČOVJEK. — Čovjek je najsavršeniji stvor među živim bićima na zemlji: on je na vrhu ljestvice kao prvak i vladar životinjskog carstva. Razlike između čovjeka i njemu najsličnijih životinja primata (majmuna i polumajmuna) s obzirom na tijelo i dušu veoma su velike. Engleski antropolog Keith navodi 312 ustrojstvenih osobina, koje su svojstvene isključivo čovjeku. Duševne su razlike još očitije: čovjeka resi govor kao manifestacija misli, etika i ostala kultura kao plod raznovrsne djelatnosti ljudskog duha, do čega se nije podigla nijedna životinja. Zato se č. mora izdvojiti iz zoologije, te po izgradnji tijela postaje unutar bioloških znanosti predmet napose antropologije (v.), a po duševnim sposobnostima predmet prehistorije, etnologije, lingvistike i psihologije.A. G-s.
SADRŽAJ: Čovjek u biološkim znanostima: A. Položaj čovjeka u živoj prirodi 335; B. Građa ljudskog tijela 335; C. Podrijetlo čovjeka, 335; D. Čovječje rase, 346; — Čovjek u prehistoriji, 366; Čovjek kao duhovno i duševno biće, 375.
Čovjek u biološkim znanostima.
A. Položaj čovjeka u živoj prirodi.
Čovjek (Homo sapiens L.) ima s obzirom na građu tijela, na fiziološke funkcije i kemijske sastojine u sistemu živih bića svoje mjesto u carstvu životinja. Već je Linné uvrstio čovjeka među sisavce, i to, zajedno s majmunima i još nekim grupama, u red Primates (prvaci). Pripisujući naročitu važnost oblikovanju krajnjih dijelova ekstremiteta Cuvier je svrstao čovjeka u zaseban red Bimana (dvorukci), dok je majmunima odredio mjesto u redu Quadrumana (četverorukci). Istoj podjeli priklonio se i Owen. Točnija su pak anatomska iskustva pokazala, da krajnji dijelovi zadnjeg ekstremiteta majmuna nisu nikakva ruka, nego lih noga, udešena za hvatanje, građena u biti kao noga kod čovjeka. I ostale anatomske razlike između čovjeka i majmuna nisu nipošto takvog omjera, kakav je u sistematici mjerodavan za razlikovanje redova (ordo), nego odgovaraju tek obilježjima, koja su značajna za porodice (familije) istog reda (→ sistematika). Izim poredbene anatomije od naročitog je značenja za položaj čovjeka u sistemu velika sličnost kemijske građe tijela čovjeka i antropomorfnih majmuna, koju nam pokazuju serološke reakcije po Friedenthalu, Uhlenhuthu i Nuttalu. Na isto nas upućuje i gotovo jednaka osjetljivost viših majmuna za specifično ljudske bolesti, napose sifilis i trahom. Svi su ovi razlozi bili mjerodavni, da se opet prihvatio za čovjeka, majmune i polumajmune Linnéov red Primates. Točan položaj čovjeka u sistemu živih bića vidi se iz priključene tabele, iz koje proizlazi i prirodoslovna definicija čovjeka.
Sadašnji je čovjek jedna jedinstvena prirodna vrsta (species). Pokušaji, da se unutar sadašnjeg ljudskog roda razlikuje više vrsta, kako je to napose činio Haeckel, koji je podijelio ljudski rod na 12 vrsta, pokazali su se neosnovanima. Razlike između različnih ljudskih rasa nisu veće nego su razlike između rasa i varijeteta kojegod životinjske rase. No kao kriterij za razlikovanje prirodnih vrsta nipošto ne mogu dolaziti u obzir intelektualne sposobnosti ili kulturni stupanj, nego samo fizička obilježja. Uz to su sve ljudske rase među sobom neograničeno plodne. Ma da to nije apsolutan kriterij za pripadnost istoj vrsti, ipak ukazuje na zajedničko podrijetlo svih sadašnjih ljudskih rasa. Jedinstvenost ljudskoga roda potvrđuje i oblik i broj kromosoma (v.); kod svih dosele ispitanih ljudi različnih rasa nalazimo po 48 kromosoma koji su morfološki posve istovetni (vidi sliku).
Položaj čovjeka u sustavu živih bića određen je samo fizičkim obilježjima. Glavna je pak, i to golema razlika između čovjeka i ostalih organizama na duševnom polju. Duševni život uzdiže čovjeka visoko iznad ostalih živih bića, te mu određuje zasebno i najviše mjesto među živim stvorovima. Svijesno svrhovito udešavanje svih pothvata, od kojega ima i kod najviših životinja tek skromnih tragova, a koje je dovelo do toga, da čovjek sebi prilagođuje prirodu, te se time rješava potrebe prilagođivanja prirodi, pojmovno mišljenje i izražavanje misli artikuliranim govorom, smisao za moralne i estetske vrednote, to su obilježja, koja određuju čovjeku njegovo vlastito duhovno carstvo.B. Z-k.
B. Građa ljudskog tijela → Anatomija, kao i članke o pojedinim ondje navedenim organskim sustavima čovječjeg tijela.U.
C. Podrijetlo čovjeka.
I. OPĆE SMJERNICE PROBLEMA.
Pitanje o postanku čovjeka kao pripadnika fizičkog svijeta dopušta dvije mogućnosti: ili postoji čovjek na Zemlji već oduvijek, ili je čovjek nastao jednom u toku povijesti Zemlje. Prva je mogućnost neodrživa, jer je gotovo sigurno, da je Zemlja u davnini bila žarko žitka, a život je bio moguć tek, kada se toliko ohladila, da se na površini pojavila voda tekućica. Uz to dosele nisu poznati fosilni tragovi čovjeka sve do najzadnjeg geološkog doba, do kvaternara. Ostaje samo druga mogućnost, da je čovjek nastao u toku razvitka žive prirode, i to u najzadnjoj fazi tog razvitka.
Tu se opet otvaraju dvije mogućnosti: ili je čovječje tijelo nastalo u biti takvo, kakvo je danas, kao zaseban stvor, neovisno od ostale žive prirode, ili se razvilo u zajednici s cijelim živim svijetom postepenim usavršavanjem u smislu teorije descendencije.
Neovisan postanak čovjeka dao bi očekivati, da je ljudsko tijelo građeno u potpunom skladu sa životnim funkcijama i potrebama čovjeka, te da je razvitak čovječjeg zametka tako udešen, da se u tom razvitku izravnim i najkraćim putem izgrađuje čovječje tijelo; isto bismo morali očekivati, da fosilni ostatci čovjeka pokazuju ista bitna obilježja, kao što ih ima i sadašnji čovjek. Treba dakle ispitati, odgovaraju li podatci anatomije, embriologije i paleoantropologije tim očekivanjima.
II. GRAĐA LJUDSKOG TIJELA KAO INDICIJ ZA NJEGOVO PODRIJETLO.
Točnije promatranje građe ljudskog tijela otkriva različne tragove, kao da naše tijelo nije neka tvorevina, izgrađena samo za sadašnje njegove funkcije i funkcije njegovih organa, nego kao da je na našem tijelu koješta preudešavano, kao kod neke građevine, koja je bila prvobitno drugojačije zamišljena, pa je za vrijeme gradnje preuređivana i pregrađivana, kako pokazuju slijedeći primjeri.
Jedan od najvažnijih organa, naše oko, i to u najhitnijem svom dijelu, mrežnici, građeno je tako, da osjetni dijelovi, štapići i čunjići, gledaju prema vani, pa da svjetlost, umjesto da izravno pogađa osjetne elemente, mora prolaziti, dok dopre do tih elemenata, kroz devet slojeva, koji kvare i svjetlost nešto raspršuju. Ako uvažimo građu jednostavnog svjetlosnog osjetila kod Amphioxusa, tada se čini, kao da je i naše oko bilo nekoć ovako građeno, pa je poslije toga dalje u tom smislu izgrađivano, i da je odatle sadašnji nezgodni položaj osjetilnih receptora u mrežnici. Kao da bi se htjelo toj neprilici u građi oka donekle doskočiti, na jednom mjestu, u središnjoj udubini u mjestu najoštrijeg vida, slojevi su razmaknuti tako, da bar na tom jednom mjestu svjetlosne zrake izravno dolaze do osjetilnih čunjića. Kao da je bila, dakle, građa oka isprva drukčije zamišljena, a zatim preudešavana.
U nutarnjem kutu oka nalazi se mali polumjesečasti nabor, koji nema nikakvog značenja. Kod nižih kralješnjaka također se nalazi ondje takav nabor, samo je vrlo prostran i služi kao treća vjeđa (membrana nictitans), koja igra izvjesnu ulogu kod zaštite oka. Čini se, kao da je i naše oko imalo prije takvu kožicu, od koje sada ima tek jedan neznatan ostatak. Druga takva tvorba bez značenja jest ono par neznatnih nabora, koji se nalaze sprijeda na tvrdom nepcu. Kod drugih sisavaca ti su nabori prošireni po većem dijelu tvrdog nepca, sa svrhom, da se na njima pod pritiskom jezika gnječi hrana. Nabori su na našem nepcu, čini se, kao neki ostatak iz prijašnjih vremena, kada su i kod naših predšasnika ti nabori imali takav zadatak. Na lopatičnoj kosti imamo kljunasti nastavak (processus coracoideus), koji ima isti položaj kao kljunasta kost kod nižih kralježnjaka. Ima slučajeva, da se taj nastavak i kod čovjeka razvija kao zasebna kost i ostaje od lopatice trajno odijeljen. Na kraju kosti krstače, gdje se kralježnica kod većine kralježnjaka nastavlja u kralješke repa, nalazimo kod čovjeka trtičnu kost, koja nema nikakvog značenja, a građena je tako, kao da je to 4—5 preudešenih i jako umanjenih repnih kralježaka, kao da je to neki ostatak prijašnjeg repa. Oko uha imamo 3 usna mišića, prednji, gornji i stražnji, koji su kod većine ljudi bez funkcije, dok su kod drugih sisavaca jako razvijeni i djelatni, te pokreću ušku. Ima različnih mišića kod drugih sisavaca, koji su ondje u punoj djelatnosti, a kod čovjeka se na istom mjestu tek katkada javljaju kao neznatni mišićni tračci bez značenja, a još češće kao vezivni tračci, koji pogotovo nemaju nikakve djelatnosti. Wiedersheim nabraja do stotinu tvorevina u našem tijelu, koje su kao neki ostatci bez prave funkcije.
S nazorom o neovisnom postanku čovječjeg tijela već je teško spojiva i sama činjenica, da je čovječje tijelo građeno po istom nacrtu kao tijelo drugih sisavaca, napose antropomorfa, da se radi samo o nešto drugojačijim omjerima pojedinih dijelova, dok te dijelove nalazimo tu i tamo u istom poretku. Ova činjenica odražava se i u položaju čovjeka u sistemu živih bića.
III. GRAĐA ZAMETKA KAO INDICIJ ZA PODRIJETLO.
Još napadnije pojave suvišnih tvorbi, koje je pogotovo teško složiti s nazorom o neovisnom postanku čovjeka, nalazimo u embrionalnom razvitku. Ma da sićušno jaje čovjeka nema gotovo nikakva žumanjka, stvara se, već prije nego se osniva tijelo zametka, žumanjčana vreća, kao da se radi o zametku, koji se razvija iz jajeta, bogato opremljenog žumanjkom. U zidu te vreće stvaraju se krvne žile, kao da bi trebale dovoditi zametku hranljive tvari iz žumanjka, kojega nema. Na tijelu zametka razvijaju se u prednjem dijelu visceralni lukovi, točno kao visceralni lukovi kod riba, od kojih se stvaraju škrge. Tada zapremaju gotovo četvrti dio tijela. U njima se stvaraju škržne krvne žile, kao da se ima razviti neko biće, koje će živjeti u vodi i disati škrgama, ali se kasnije veći dio tih lukova i njihovih žila razgrađuje. Prvobitna čeljust čovjeka stvara se iz prednjeg luka, kao kod nižih kralježnjaka; kao tu se posebna t. zv. kvadratna hrskavica povezuje s lubanjom, a da bude u kasnijem razvitku zametka veći dio te čeljusti razgrađen i zamijenjen novom čeljusti, koja je neposredno zglobovno spojena s lubanjom. U početku drugog mjeseca čovječji je zametak opremljen repom, koji kasnije zakršlja, a ostatak njegova skeleta predstavlja trtična kost. U razvitku gotovo svakog organa čovjeka možemo naći takvih primjera.
IV. BIOLOŠKO TUMAČENJE ANATOMSKIH I EMBRIOLOŠKIH INDICIJA.
Kako pod vidom neovisnog postanka čovjeka za spomenute i slične pojave u građi i razvitku čovjeka nijedno tumačenje ne zadovoljava, nego se te pojave čine protuslovnima i besmislenima, biološka se znanost sve više priklonila shvaćanju, da je čovjek kao fizičko biće nastao postepenim razvitkom iz nižih organizama u smislu descendentne teorije, da je čovjek zajedničkog podrijetla s ostalim živim bićima, i to, da se njegova loza odcijepila od debla, kojega ostale grane vode k onim sisavcima, koji su čovjeku najsličniji, majmunima. Već kod Buffona nalazimo tragove tog shvaćanja, a prvi je to jasno izrazio Lamarck (1809), i to samo na temelju sličnosti građe čovjeka i majmuna. Kad je Darwin izašao sa svojom teorijom (1859), primijenio je Th. Huxley (1863) na temelju anatomskih obilježja kao prvi tu teoriju na čovjeka. Za njim su slijedili drugi biolozi, napose E. Haeckel (1866), pa i Darwin sam, koji je u posebnom djelu (1871) sabrao tada poznate indicije za razvitak čovjeka iz nižih forma. Vulgarno se često taj nazor naziva »majmunskom teorijom«, što međutim nije točno, jer osim Lamarcka nitko nije tvrdio, da se čovjek razvio iz majmuna, nego jedino, da je čovjek zajedničkog podrijetla s ostalim primatima, da su čovjek i drugi sadašnji primati vršci razvojnih grana, koje su se u davnini odvojile od zajedničkog debla. To je današnje gotovo jednodušno shvaćanje predstavnika različnih grana biološke znanosti; različita mišljenja postoje jedino u tome, kada i na kojem stupnju organizacije se razvojna loza čovjeka odcijepila od zajedničkog debla.
Dok poredbena anatomija i embriologija pružaju indicije tek za glavne smjernice podrijetla čovjeka kao fizičkog bića, to su za pitanje, koji su bili zadnji njegovi predšasnici, i gdje i kada se odcijepila njegova loza, od najveće važnosti fosilni ostatci, koji uz to pružaju daljne indicije za razvitak čovjeka u smislu transformizma. Dok je u prošlom stoljeću bilo poznato još malo takvih ostataka, tako da im je R. Virchow mogao poricati svako značenje, te je veliku ulogu igralo traganje za »missing-link«-om (karikom, koje nema, naime u lancu razvojnih stupnjeva), to je uspjelo pronaći cijeli niz fosilnih hominida, pošto je Dubois otkrio okamine na Javi, a Gorjanović-Kramberger svojim čuvenim nalazom u Krapini postavio nauku o pračovjeku na čvrste temelje, i otada gotovu svaka godina donaša nova važna otkrića.
V. FOSILNI OSTATCI HOMINIDA.
Između fosilnih ostataka hominida, različnih od sadašnjeg čovjeka, ima tipova, koji se radi svojih primitivnih obilježja pribrajaju posebnim rodovima. Naziv tih rodova svršava se riječju — anthropus, zato se zovu skupinom Anthropusa. Dalje ima tipova, koji svakako pripadaju rodu Homo, ali se smatraju posebnim vrstama, među kojima možemo donekle razlikovati primitivniju predneandertalsku te neandertalsku skupinu. Napokon, ima ostataka pravog čovjeka, koji pripadaju drugim rasama nego su sadašnje; tu skupinu nazivaju Homo sapiens fossilis. Najvažnije od tih dosele poznatih fosilija bile bi u morfološkom redosljedu slijedeće:
a) Anthropus-skupina.
Pithecanthropus erectus E. Dubois. (πίϑηϰος »majmun«, ἄνϑρωπος »čovjek«, erectus »uspravan«). Tab. sl. 3—6.
Pronašao ga je 1891/92 E. Dubois kod Trinila na Javi, i to najprije 1 kutnjak, nato kalotu (gornji dio moždane lubanje), još 1 kutnjak, a 15 m dalje lijevu bedrenjaču (femur). Kasnije je Dubois među svojedobno sakupljenim okaminama pronašao još komadić donje čeljusti i komade od 4 bedrenjače. Poslije 40-godišnjeg traganja različnih istraživača uspjelo je 1937 Koenigswaldu u istom ležištu kod Sangirana pronaći još jednu kalotu uz velik komad donje čeljusti, a 1938 kod Kedung-Brubusa isti je istraživač pronašao još komade treće lubanje. Ležište se sastoji iz vulkanskog tufa i nalazi se neposredno na morskim pliocenskim naslagama. Stoga je Dubois držao, da ležište pripada gornjem pliocenu, ali se kasnije ispostavilo, da je mlađe. Van Es drži, da pripada srednjem diluviju. Do istog je rezultata nakon pomnih istraživanja došao Koenigswald (1939), naime, da trinilsko ležište vjerojatno potječe iz interglacijalnog doba Mindel-Riss.
Kalota pokazuje jake nadočne nabubrine (torus supraorbitalis), iza očne šupljine jako je sužena, što sjeća na antropomorfe. U istom se smislu ističe brid na zatiljnoj kosti i jako plitko čelo. Prva kalota, koja potječe od muškog bića, odgovara lubanji sa sadržajem od kojih 900 cm3, novo pronađena, vjerojatno ženska, sa sadržinom od 750 cm3 (gornja granica kod gorile 600 cm3, donja granica kod čovjeka — žene australske rase — 950 cm3, kod Evropejca je srednja vrijednost 1300 cm3). Odljevak lubanje pokazuje, da je treći česni zavoj mozga, Broca-ov zavoj govora, imao samo polovicu opsega kao kod sadašnjeg čovjeka. Čeljust je masivna, za ⅓ jača nego kod sadašnjeg čovjeka, jednako su kutnjaci veliki i široki, zadnji kutnjak još ne pokazuje nikakve redukcije. Bedrenjače pak gotovo su jednake kao kod sadašnjeg čovjeka, tako da je izražena sumnja, pripadaju li istoj vrsti kao i kalota, pogotovu, jer je najbliža nađena 15 m dalje od kalote. Ali kako je nađena u istom sloju, a fosilizacija je točno ista kao kod kalote, opravdava se time mnijenje, da je iste vrste, dapače, od istog individuuma, ma da se u zadnje vrijeme opet sve više javlja shvaćanje, da su to bedrenjače od drugovrsnog, čovjeku sličnijeg bića.
U početku je taj nalaz različito prosuđivan. R. Virchow, Krause, Waldeyer, Ranke, Sergi, R. Martin tvrdili su, da je kalota od velikog antropomorfnog majmuna, a bedrenjača od čovjeka; Topinard, Houzé, Lydekker, Turner, Cunningham, da se radi o primitivnom čovjeku; većina stručnjaka, Dubois, Verneau, Manouvrier, Nehring, Schwalbe, Haeckel, Klaatsch, Duckworth, Sollas, Keith, Marsh, Osborn i dr., da je to »missing-link«, prelazna forma između predšasnika antropomorfa i čovjeka. Dubois je tvrdio, da ima kalota mnogo sličnosti s gibonom, ali je kasnije Weinert upozorio, da postoji šupljina u čeonoj kosti (sinus frontalis), koje gibon nema, pa da je Pithecanthropus morfološki bliži čimpanzi.
Treća lubanja pokazuje mnogo sličnosti sa Sinanthropus-om. Čelo, naime, nije tako plitko kao kod prve dvije lubanje, tako da se time potvrđuje, da su postojale bliske veze između javanskog i pekinškog čovječnjaka.
Sinanthropus pekinensis D. Black. (Sina=Kina). Tab. sl. 7—9.
Pošto je 1923 Zdanski pronašao kod Šu-ku-tjena kraj Pekinga jedan zub, nalik na čovječji, započeo je 1927 Dawidson Black s iskopavanjima. Pronađen je najprije zub, 1928 donja čeljust mladica, komadi čeljusti odrasloga i donji zubi, a 1929 naišao je Pei na moždani dio lubanje i na različite zube; g. 1930 nađeni su komadi lubanje, iz kojih se dala sastaviti još jedna kalota. G. 1931 pronađena su u susjednoj Kot-dze-tang-spilji 2 komada čeljusti i komadi lubanje uz tragove garišta, a 1932 oruđe iz kamena i komadi paočnice (radius). Poslije B1ackove smrti nastavio je s iskopavanjima Weidenreich i naišao na ostatke od kojih 28 individuuma, među njima komade, iz kojih je uspjelo sastaviti 3 daljnje lubanje. Nađeni su i komadi kosti ekstremiteta. Taj materijal još nije do kraja obrađen. Ležište kod Šu-ku-tjena sastoji se iz naslaga u nekoj spilji, kojoj se svod srušio. Svi su slojevi zajedno debeli do 50 m. U najnižem sloju pronađena je prva lubanja, a ostale kosti u svim slojevima. Tu pronađeni ostatci različnih sisavaca pokazuju, da naslage potječu iz doba vlažne i umjerene klime, pa se zaključuje, da je ležište iz Mindel-Riss-interglacijalnog doba, dok ga Pei (1939) drži starijim, naime iz interglacijala Günz-Mindel.
Ovo, dosele najznatnije paleoantropološko otkriće u 20. st., pokazuje nam vrstu homínida, koja je vrlo bliska Pitheanthropus-u, te bi bilo opravdano i pekinški nalaz prozvati tim imenom. Razlika se sastoji u tome, što čelo nije tako plitko, nego lako izbočeno iznad uvlake, koja se nalazi iza jakih nadočnih nabubrina. Sljepočni je šav gotovo ravan kao kod antropomorfa, zatiljna kost ima jaki brid. Gledana odozgo, lubanja iza očne šupljine nije tako jako sužena kao kod Pithecanthropus-a. Sadržina lubanja koleba između 850 i 1220 cm3. Doljnja je čeljust masivna, bez bradnog šiljka, ali je, u poredbi sa čeljusti čimpanze ili gorile, jako kratka. Prema čovječjoj čeljusti od strane gledana izvanredno je visoka, što joj daje neobičan izgled. Zubi su čvrsti i veliki te pokazuju u nekim obilježjima mnogo sličnosti sa zubima antropomorfa, napose čimpanze, ali ima i svih prijelaza k zubima sadašnjeg čovjeka. Kasnije pronađene lubanje su na tjemenu nešto izbočenije, te se donekle približuju neandertalskom pračovjeku.
Nakon nalaza prve lubanje Sera je držao, da nije normalna, jer pokazuje t. zv. platibaziju (splošteno dno); no ovaj je prigovor oboren daljnjim nalazima jednakih lubanja. Dubois je izrazio mišljenje, da Sinanthropus ide u varijacioni opseg neandertalca, a Sera je našao sličnosti napose s gibraltarskom lubanjom. Nalazi pak daljnjih lubanja pokazuju, da je to bio zaseban, jasno ograničen tip, koji ima najbliže veze s Pithecanthropus-om, ali već čini morfološki prijelaz k pračovjeku. Weidenreichu je uspjelo pronaći i lične dijelove lubanje, i to ženske, tako da je mogao s pomoću tih komada sastaviti cijelu žensku lubanju Sinanthropus-a. Iz te restauracije lubanje proizlazi, da je bila jako kosozuba (prognatna), mnogo jače, nego kod ikoje sadašnje ljudske rase. Kosti ruke i noge, od kojih je najveća 31 cm dugi komad bedrenjače, tipično su čovječje, bez naročitih sličnosti s udovima antropomorfa.
Od osobite je važnosti činjenica, da su u istom ležištu pronađeni tragovi garišta i kameno oruđe, iz čega slijedi, da su Sinanthropus, a prema tome i njemu tako slični Pithecanthropus, bili inteligentna bića, koja su poznavala priređivanje vatre i proizvodila i upotrebljavala kameno oruđe. Time je pogotovo obeskrijepljeno mnijenje, da lubanja Pithecanthropus-a pripada velikom majmunu, a ujedno je to daljnji indicij, da bi femur iz Trinila mogao doista pripadati Pithecanthropus-u, jer samo biće, koje uspravno hoda, može se služiti rukama, da sebi priređuje oruđe.
Nalaz oruđa i tragova garišta pokušao se, doduše, i tako tumačiti, da nije Sinanthropus palio vatru i služio se oruđem, nego neki viši pračovjek-ljudožder, koji je Sinanthropus-e ubijao i pržio na vatri, pa da su kosti Sinanthropus-a ostatci tih gozba. Ali bi čudno bilo, a nema nikakvog traga od kosti tog ljudoždera. Uz to bi se morale baš onda naći i različne druge kosti Sinanthropus-a, a pronađene su gotovo samo lubanje. To se shvaća tako, da je Sinanthropus, zbog nekih mističnih predodžbi, običavao nositi sa sobom lubanje umrlih drugova, kako to još danas čine urođenici na Andamanskim otocima.
Africanthropus njarassensis Weinert. Tab. sl. 10.
G. 1935 našao je etnolog Kohll-Larsen kod Njarasa-jezera u istočnoj Africi komade jako mineraliziranih lubanja. Komadi su virili iz pješčenjaka, očito razgrađenog djelovanjem vjetra. Komadi potječu od 2 ili 3 lubanje. Starost se nije dala točno odrediti, a to vrijedi za većinu afričkih nalaza fosilnog čovjeka. Fauna se onog ležišta sastoji iz sadašnjih i nekih izumrlih forma. Reck drži, da se radi o gornjem diluviju, dok Weinert tom nalazu daje mnogo veću starost, no jedino s obzirom na mineralizaciju. Po različnim iskustvima u tropskoj Africi kosti se brzo mineraliziraju. Svakako pitanje starosti nije razjašnjeno i valjda se ne će moći nikad sigurno riješiti.
U koliko je uspjelo komade sastaviti, što su najprije pokušali Reck i Leakey, a nato Weinert, predočuju nam moždani dio lubanje, vrlo slične Pithecanthropus-u, samo što zatiljna kost nema tako izrazit brid, nego je lako zaobljena. Sadržina lubanje iznosila bi oko 1100 cm3.
Eoanthropus dawsoni Smith-Woodward. Tab. sl. 16 i 17.
G. 1909 iskopali su seljaci na jednoj njivi kod Piltdowna (Sussex) u Engleskoj okamenjenu lubanju, koja je kod toga razbijena, i bacili te komade na gomilu šljunka. Geolog Dawson, koji je saznao za to, pretražio je ovaj šljunak i dalje kopao, pa su pronađeni još neki ostatci, a 3 km dalje u jednakom ležištu donja čeljust, od koje nema prednjeg dijela. Fauna, pronađena u tim naslagama, odgovara pliocenu ili najstarijem diluviju. Nije pak sigurno, nisu li ostatci one faune već kao fosili došli u to ležište, pa da je samo ležište mlađeg datuma.
Smith-Woodward sastavio je iz pronađenih komada lubanju, koja u biti pokazuje obilježja pravog čovjeka. Sadržinu je procijenio na 1070 cm3. Keith je lubanju na novo sastavio, a sadržina bi joj prema toj rekonstrukciji bila 1377 cm3, dakle kao kod sadašnjeg čovjeka.
Dok je lubanja gotovo ljudska, donja je čeljust izvanredno slična čeljusti čimpanze, samo je veća. Ako se k tome uzme u obzir, da je nađena 3 km daleko od lubanje, lako se nameće tumačenje, da lubanja i čeljust ne spadaju zajedno. Stoga su mnogi stručnjaci, među njima G. S. Mi11er, M. Boule, Runnström, Werth, mnijenja, da je lubanja od pravog čovjeka, a čeljust od velike vrste čimpanze, koja je tada živjela u Engleskoj, pa su je nazvali Pan vetus. Fredericks poređuje čeljust s onom od oranga. No svi engleski stručnjaci, koji su ostatke proučavali u originalu, među njima i vrlo kritični Keith, drže, da su lubanja i čeljust od istog bića. I Hrd1ička, koji je 1925 ispitao originalne kosti, priključuje se tom mišljenju. Weinert, koji je isprva bio prihvatio mišljenje o dvostrukom podrijetlu čeljusti i lubanje, također se izrazio za jedinstvenost, kad je proučio originalne kosti. Po svemu je, dakle, ipak vjerojatnije, da se tu radi o izumrlom hominidu svoje vrste, ali koji s obzirom na donju čeljust ne bi spadao u izravnu razvojnu lozu čovjeka, nego bi zastupao neku postranu granu.
b) Preneandertalska skupina.
Homo Heidelbergensis Schoetensack. Tab. sl. 14.
G. 1907 iskopana je u kamenolomu blizu sela Mauer kod Heidelberga vrlo dobro sačuvana fosilna donja čeljust, koju je Schötensack svestrano proučio. Ležište se nalazi 25 m ispod površine u naslagama, koje potječu iz interglacijalnog doba Günz-Mindel.
Stražnji je dio (ogranak) ove donje čeljusti vrlo širok i jak kao kod antropomorfa, dok trupac više sjeća na ljudsku čeljust. Zubi su, premda vrlo jaki i masivni, tipično čovječji, očnjak ništa ne iskače iznad ostalih zubi. Na žalost, postoji samo taj ostatak, po kojem ne možemo zaključiti, kakva je bila ostala lubanja.
Steinheimska lubanja.
Iskopao ju je 1933 kod Steinheima u Württembergu F. Berckheimer, a obradio napose H. Weinert. Ležište potječe vjerojatno iz toplije periode, koja se javila za vrijeme Risskog ledenog doba.
Radi se o ženskoj lubanji, koja nema s lijeve strane dijela ličnih kosti. Čelo je plitko, ali ipak nešto izbočeno, tako da se ta lubanja nadovezuje na Sinanthropusa. Zatiljna je kost u profilu zaobljena. Lične pak kosti sjećaju u mnogome na sadašnjeg čovjeka, tako da bi se od Steinheimskog čovjeka mogla izvesti izravna loza, koja vodi k sadašnjemu. Pogotovu je dobilo ovo shvaćanje oslona u građi lubanje, pronađene kod Swanscombea, koja ima mnogo sličnosti sa steinheimskom, ma da je već savršenija. Sadržina Steinheimske lubanje iznosi oko 1070 cm3.
Homo rhodesiensis Smith-Woodward. Tab. sl. 11. i 12.
U rudniku kod Broken-Hilla u Rodeziji (Juž. Afrika) iskopana je 1921 lubanja odraslog muškarca, a 1 m dalje lijeva cjevanica (tibia) i komadi različnih kosti, valjda od više osoba. Ležište sadržava faunu, koja se od sadašnje afričke faune gotovo ne razlikuje, tako da se starost ležišta, kao kod većine afričkih kvarternarnih ležišta, ne može sigurno odrediti. Svakako, nije starije od gornjeg diluvija, a možda je i mlađe. Za potonje govori činjenica, da zubi pokazuju tragove karijesa (gnjiloće), a zubni karijes javlja se tek u novije doba.
Na ovoj lubanji, koju je obradio Smith-Woodward, zapanjuje plitko čelo i upravo goleme nadočne nabubrine, najjače od svih lubanja fosilnih hominida. Zatiljna kost ima oštar brid. Sadržina je procijenjena na 1300 cm3, dakle je relativno velika. Odljevak moždane šupljine pokazuje, da su čeoni i sljepočni režanj mozga bili na vrlo primitivnom stupnju. Rodezijskog čovječnjaka možemo shvatiti kao zadnji vršak neke postrane loze, koja se u Africi sačuvala još do vremena, kada je u ostalom svijetu već davno preuzeo vlast sadašnji čovjek.
Homo modjokertensis Koenigswald. Tab. sl. 15.
G. 1936 iskopan je kod Modjokerte na Javi gornji dio lubanje 1½ godišnjeg djeteta. Ležište je iz donjeg diluvija, dakle starije od ležišta Pithecanthropus-a.
Dok je kod dječje lubanje sadašnjeg čovjeka čelo okomito ili naprijed izbočeno, ova lubanja ima dosta nagnuto čelo, donekle nalik na čelo dječje lubanje fosilnog antropomorfa Australopithecus-a. Sadržina iznosi 730 cm3, što bi odgovaralo sadržini lubanje odraslog od 1100 cm3. Svakako se radi o biću, koje je bilo na višem stupnju nego Pithecanthropus, pa se ta lubanja može najispravnije pripisati čovječnjaku predneandertalske skupine.
c) Neandertalska skupina.
Homo neanderthalensis King, pračovjek ili neandertalski čovjek. Tab. sl. 18—23.
Svoje ime nosi ta forma od čuvenog nalaza u spilji u Neanderthalu kod Düsseldorfa, gdje su 1856 radnici našli kalotu lubanje i brojne dijelove okosnice, koje su opisali Fuhlrott i Schaffhausen. Daljnji su pronalasci pračovjeka slijedeći:
| God. |
Nalazište |
Predmet |
Autor |
| 1848 |
Gibraltar |
lubanja |
Busk (opisao 1868) |
| 1865 |
La Naulette (Belgija) |
donja čeljust |
Dupont |
| 1882 |
Šipka (Moravska) |
donja čeljust |
Maška |
| 1886 |
Spy (Belgija) |
2 okosnice |
Fraipont |
| 1887 |
Bañolas (Španjolska) |
donja čeljust |
Pocheco i Obermaier (1915) |
| 1889 |
Malarnaud(Francuska) |
donja čeljust |
Filhol |
| 1895 |
Taubach (Njemačka) |
2 zuba |
Nehring |
| 1899 |
Krapina (Hrvatska) |
brojne okosnice |
GorjanovicKramberger |
| 1906 |
Ochoz (Moravska) |
donja čeljust |
Rzehak |
| 1908 |
Le Moustier (Francuska) |
okosnica |
Klaatsch i Hauser |
| 1908 |
La Chapelle-aux-Saints (Francuska) |
okosnica |
Boule |
| 1909 |
La Ferassie (Francuska) |
2 okosnice |
Peyrony |
| 1909 |
Pech de l’Azé (Francuska) |
dječja lubanja |
Peyrony |
| 1910 |
St. Brélade (Francuska) |
zubi |
Keith |
| 1912 |
La Quina (Francuska) |
okosnica |
H. Martin |
| 1914 |
Ehringsdorf (Njemačka) |
donja čeljust |
Schwalbe |
| 1916 |
Ehringsdorf (Njemačka) |
donja čeljust |
H. Virchow |
| 1925 |
Ehringsdorf (Njemačka) |
lubanja u komadima |
Weidenreich |
| 1925 |
Tabgha (Galileja) |
dio lubanje |
Turville-Petre |
| 1926 |
Gibraltar |
dječja lubanja |
Carrow |
| 1929 |
Saccopastore (Italija) |
2 lubanje |
Sergi |
| 1931 |
Kiik-Koba (Krim, Rusija) |
2 okosnice |
Bonc-Osmolovskij |
| 1931/32 |
Sukul (Palestina) Tabun (Palestina) Džebel Kafze (Palestina) |
10 okosnica čeljust i ženska okosnica 5 okosnica |
Garrod, Mac Cown i Neuville |
| 1939 |
Monte Circeo (Italija) |
lubanja |
Blanc |
Ležišta pračovjeka popraćena su faunom, koja odgovara toploj klimi zadnjeg interglacijalnog doba (Riss-Würm), ili faunom, koja odgovara već hladnijoj klimi, dakle početku zadnjeg (würmskog) ledenog doba, što znači, da je pračovjek vladao zemljom više od 100.000 godina.
Lubanja pračovjeka pokazuje vrlo masivne kosti, tako da je R. Virchow, s obzirom na nepotpune nalaze u Neanderthalu i Spyju, tvrdio, da su to bolesno pretvorene kosti. Kasniji su nalazi, počevši s Gorjanovićevima u Krapini, pokazali, da je taj »bolesnik« bio svojevrsna forma čovjeka, proširena od Španjolske sve do južne Rusije i Palestine. Glavna su obilježja pračovjeka: Tijelo niskog uzrasta s masivnim jakim kostima. Glava velika. Lični dio lubanje mnogo jače razvijen nego kod sadašnjeg čovjeka. Donja čeljust sprijeda koso odsječena bez bradnog šiljka, njezin stražnji dio širok. Nosni otvor vrlo širok. Očne šupljine upravo goleme, a nad njima jake nadočne nabubrine. Čelo plitko. Moždani dio lubanje splošten, tako da ima zatiljna kost jako kos položaj. Noge relativno kratke. Bedrenjača (femur) ukrivljena. Trnoviti nastavci na vratnim kralješcima stoje gotovo vodoravno, ne koso, kao kod nas. Uz to još cijeli niz sitnijih obilježja, koja sjećaju na prilike kod antropomorfa.
Sadržina lubanje prilično je velika, prosječno 1450 cm3, dakle nipošto manja nego kod sadašnjeg čovjeka. Kod toga treba naravno uvažiti, da za prosuđivanje kakvoće mozga sadržina lubanje ne znači baš mnogo. Površina moždane kore ovisi prije svega o razbrazdanosti mozga. Neki se zavoji mozga mogu još donekle razabrati na odljevcima nutrine lubanje. Od najhitnijeg pak značenja po kakvoću mozga jest gustoća i poredak stanica u kori, t. zv. citoarhitektonika, o kojoj nam sadržina lubanje ne može ništa kazati. Odljevci moždane lubanje pračovjeka u ostalom pokazuju, da su moždani zavoji bili primitivni i široki, napose je treći čeoni zavoj, gdje se nalazi središte govora, bio dosta jednostavan. Boule drži, da pračovjek nije hodao sasvim uspravno, nego, radi položaja trnovitih nastavaka, s prignutom šijom, dok Weidenreich to s obzirom na građu kukovlja poriče. Kameno i koštano oruđe i tragovi garišta, koji popraćuju nalaze kosti pračovjeka, pokazuju, da je pračovjek znao sve to priređivati i upotrebljavati.
Ostatci pračovjeka s različitih nalazišta pokazuju izvjesne razlike. Tako se gibraltarska lubanja ističe po nekim obilježjima, upravo golemim otvorom nosne šupljine i jakom sploštenošću moždanog dijela. Na nju podsjeća i lubanja iz Saccopastore. Čeljust iz Ehringsdorfa na sredini (simfizi) je jako naprijed izbočena. Napose pokazuju veliku varijabilnost nalazi iz Palestine. Tu su razlikovali tabunski tip s obilježjima tipičnog neandertalca i sukulski tip, koji je izglađeniji i sjeća na fosilnog čovjeka iz Cro-Magnona. Donja čeljust ima kod potonjeg već kao neki početak bradnog šiljka. Ima pak i forma, koje ona dva tipa povezuju. M. F. Ash1ey-Montagu dolazi stoga do zaključka, da bi se moglo raditi djelomično o križancima između neandertalca i pravog čovjeka, dok drugi u onim posrednim tipovima vide prijelaze između neandertalca i najstarijih forma pravog čovjeka. Svakako ta raznolikost pokazuje, da je pračovjek bio pocijepan na rase, pa da je i inače pokazivao varijabilnost, što je i s obzirom na razdoblje bar od kojih 100.000 godina njegova opstanka razumljivo. Kao neka oštrije odijeljena rasa pračovjeka ili kao zasebna vrsta može se smatrati
Homo soloensis Oppenoorth, nazvan i Javanthropus, pračovjek s otoka Jave. Tab. sl. 24.
Uz rijeku Solo, uz koju se nalazi i Trinil, nalazište Pithecanthropus-a, i to 10 km od tog nalazišta, kod Ngandonga, iskopao je 1932 Oppenoorth brojne ostatke od 11 lubanja, od kojih je 6 sačuvano kao kalote. Ležište je diluvijalno, i to vjerojatno iz Riss-Würmskog interglacijalnog doba.
Lubanje pokazuju uglavnom obilježja Neandertalca, ali se sve ističu naročitom plitkošću čela i na zatiljku osobito velikim pričvršćenim poljem mišića sije. Sadržina im se kreće između 1200 i 1300 cm3.
d) Skupina Homo sapiens fossilis. Tab. sl. 25—30.
Iz naslaga zadnjeg (würmskog) ledenog doba, te napose iz prapora (Löss), koji su vjetrovi nanosili u stepe onog hladnog vremena, ima brojnih ostataka pravog čovjeka. Sob, mamut, dlakavi nosorožac, spiljski medvjed bili su njegovi suvremenici. Među tim čovječjim fosilijama ima tipova, koji se vrlo približuju obilježjima izvjesnih rasa sadašnjeg čovjeka, a ima i tipova, koji sjećaju još na pračovjeka. Svima tim tipovima diluvijalnog čovjeka dao je Schwalbe ime Homo sapiens fossilis. Svakako je i diluvijalni čovjek bio pocijepan na rase, ali su svi ti tipovi povezani prelaznim likovima, tako da se mišljenja razilaze o tome, možemo li ograničiti pojedine rase.
Klaatsch i Hauser su na osnovi okosnice, pronađene 1909 u spilji Combe Capelle u južnoj Francuskoj, ovaj tip zajedno s nekim sličnim tipovima proglasili posebnom Aurignacskom rasom.
Istoj grupi pribrajaju još slijedeće nalaze: okosnicu iskopanu 1891 u Brnu; brojne nalaze iz Předmostí u Moravskoj, gdje su iskopavali od 1882 dalje Maška i na to Křiž, i to okosnice od 8 odraslih i 12 mlađih osoba; kalotu iz Brüxa od 1871; lubanju, koju je 1883 iskopao Frič u praporu u Podbabi kod Praga; napokon ima slična obilježja i lubanja od Galley-Hilla na ušću Temze, koju je iskopao Elliot 1888. Za potonju lubanju, doduše, nije sigurno, nije li tek naknadno došla u one diluvijalne naslage.
Čelo je kod tih tipova još dosta plitko, lubanja uska i dugačka, zatiljak već zaokružen. Lični je dio uzak i visok. Donja čeljust kod nekih lubanja ne pokazuje još jasan šiljak na bradi, nego je brada ravno odsječena. Ostala okosnica mnogo je gracilnija nego ona u pračovjeka. Klaatsch nalazi izvjesne sličnosti sa sadašnjom australskom rasom.
Iz gornjeg diluvija, napose iz doba, kada su se već počeli würmski ledenjaci povlačiti, potječu okosnice s obilježjima, koja bi se mogla shvatiti kao odlike naročite t. zv. Cro-Magnonske rase.
Ovamo idu okosnice, koje je 1868 Lartet iskopao u Cro-Magnonu, u dolini rijeke Vezère; dalje okosnica, koju je 1872 kod Laugerie-Basse u istoj dolini pronašao Massénat; okosnica, koju su 1888 našli Féaux i Hardy kod Chancelade; ostatci iz Duruthya i Les Hoteauxa; okosnice iz gornjih slojeva spilje kod Mentonea, gdje su od 1872 do 1892 kopali redom Rivière, Julien, Abbo i napokon Verneau; 2 okosnice, muška i ženska, pronađene 1914 kod Oberkassela kod Bonna i obrađene od Verworna, Bonneta i Steinmanna i dr.
Cro-Magnonski tip vrlo je robusne građe, visokog stasa (175—189 cm), lubanja je dugačka, ali je lični dio širok i kratak sa širokim očnim šupljinama. Donja čeljust ima izrazitu bradu. Taj tip nalazimo u bitnim njegovim potezima još danas kod stanovnika Kanarskih otoka i kod predstavnika dalijske rase.
Za nalaz od Chancelade istaknuo je već Testut (1890), da obilježja lubanje potsjećaju na eskimske lubanje. Visinu tijela je procijenio po duljini bedrenjače na 159 cm, što bi se podudaralo sa stasom Eskima. Zato drže neki stručnjaci, među njima i Eickstedt, da se radi o posebnoj rasi, pogotovu, jer su 1928 pronađene kod Le Roc-a 3 slične okosnice.
Oštrije se razlikuje od spomenutih tipova Grimaldi-rasa. Radi se o 2 okosnice, od mlađeg muškarca i starije žene, koje je 1901 knez Albert od Monaka iskopao u najdonjem sloju naslaga u spilji »Grotte des enfants« kod Mentonea, a obradio ih je R. Verneau.
Lubanje su uske i dugačke, kosozube (prognatne), pa u tome kao i nekim obilježjima pokazuju mnogo sličnosti s lubanjom današnjih Crnaca. Stas je bio nizak, 156—159 cm. Noge su jako dugačke zbog relativno velike duljine golijeni, a jednako je i podlaktica relativno dugačka, što sve također sjeća na Crnce, a još više na Negrite. Ipak time nije kazano, da bi bila ova rasa predšastnik Crnaca. Verneau drži, da taj tip predočuje primitivnu formu Cro-Magnonaca.
Vadjak. Tab. sl. 31. U diluvijalnom ležištu kod Vadjaka na Javi iskopao je 1886 E. Dubois komade čeljusti i lubanju, koja pokazuje plosnato čelo s jedva naznačenim izbočenjem, svojim pak širokim očnim šupljinama podsjeća na Cro-Magnonce. Dubois je taj tip nazvao protoaustralskim. Weinert drži, da je to daljni razvojni stupanj pračovjeka iz Ngandonga i predšasnik sadašnjih Australaca.
Kanam i Kanjera. Od naročitog značenja za pitanje, kada je nastao pravi čovjek, Homo sapiens, mogao bi biti nalaz u Kanamu kod Viktorijina jezera u središnjoj Africi, gdje je 1931 L. S. B. Leakey pronašao prednji dio ljudske čeljusti s tipičnim bradnim šiljkom, dakle tipične čeljusti vrste Homo sapiens, ali u ležištu, u kojem su pronađeni ostatci izumrlih slonova Dinotherium i Mastodon, što bi odgovaralo donjem diluviju. Ležište sastoji iz naslaga Viktorijina jezera, kojega je razina u ono doba bila mnogo viša nego danas. To je ležište mnogo starije nego ležišta Pithecanthropus-a ili neandertalca. Nešto je mlađe, iz srednjeg diluvija, ležište u Kanjeri, gdje je Leakey pronašao ostatke čovječje lubanje. Komisija brojnih stručnjaka, koja je u Londonu 1933 ispitala te okamine, izjavila se za njihovu visoku starost, kao i za to, da pripadaju vrsti Homo sapiens. Geolog Boswell, koji je poslije toga ispitao ona ležišta u središnjoj Africi, izrazio se vrlo skeptički i ne vjeruje u tako veliku starost tih ostataka. Leakey pak i nakon toga ostaje kod svoje tvrdnje o starosti onih fosilija.
Swanscombe. S time, da bi već u srednjem diluviju, a možda i ranije, živio pravi čovjek, slaže se i nalaz kod Swanscombea na donjoj Temzi u Engleskoj, gdje je 1935 A. T. Marston pronašao u šljunku t. zv. 100-stopne terase rijeke Temze zatiljnu kost nalik na čovječju, a godinu dana kasnije 7 m dalje tjemenu kost. Ležište potječe bez sumnje iz Mindel-Riss-interglacijalnog doba. Kosti, koje je ispitala komisija stručnjaka, spadaju jednoj lubanji, i to vjerojatno ženskoj. Sadržina čitave lubanje procijenjena je na 1325 cm3. Odljevci nutrine pokazuju dobro razvijene zavoje. Od sadašnjeg čovjeka razlikuju se te kosti po širini zatiljka, i što su vrlo debele. U profilu je zatiljak lijepo zaokružen bez ikakva brida. Svakako se dakle radi o formi, vrlo bliskoj pravom čovjeku. Mnogo zajedničkih obilježja ima taj nalaz sa Steinheimskom lubanjom, tako da bi se mogla, kako već istakosmo, odatle preko Swanscombskog tipa izvesti loza sadašnjeg čovjeka.
Na prekretnici wurmskog ledenog doba i aluvija javljaju se čovječji tipovi, na koje se već priključuju oblici današnjeg čovjeka. To su u Europi napose nalazi iz spilje Mas d’Azil u Francuskoj i spilje Ofnet u Bavarskoj, u kojoj su pronađeni ostatci od 33 okosnice. Lubanje iz spilje Ofnet pokazuju već sve oblike, koje i danas nalazimo u Evropi, ima dolihocefalnih i brahicefalnih. I na nalaze jednake starosti u drugim zemljama isto se priključuju današnji tipovi u tim zemljama. Tako su u Australiji kod Cohune, kod Jervoisa i kod Talgaja nađeni ostatci čovjeka, od kojih potječe potonji, dapače vjerojatno iz gornjeg diluvija. Njima se, doduše, pokušalo pripisivati naročito značenje, ali su se napokon ispostavili kao forme vrlo slične današnjim urođenicima tog kontinenta. Isto vrijedi za različne nalaze u Africi, među kojima je neko vrijeme bio vrlo važan nalaz iz sutjeske Oldoway u istočnoj Africi, gdje je 1913 Reck našao čovječju okosnicu, kojoj je pripisao veliku starost. Nalaz u Oldowayu sastoji se iz 5 slojeva. Donja 4 potječu iz pluvijalnog doba, koje odgovara ledenomu doba u Evropi. U 2. sloju nalazila se okosnica, ali u položaju čučanja, što znači, da je bila u tom položaju pokopana i tako došla u ono ležište. Boswell je našao u grudnom košu te okosnice zemlju iz najgornjeg sloja, što znači, da je okosnica iz doba stvaranja te površne naslage ili je još mlađa. Kako su Mollison i Gieseler pokazali, specifična je težina tih kosti ista kao svježih ljudskih kosti. Mollison je upozorio na to, da lubanja ima sličan tip, kao sadašnji Crnci plemena Masai. S time se slaže i visina tijela od 180—184 cm.
Svi nalazi fosilnih hominida iz diluvija potječu, kako je vidljivo, iz staroga svijeta. Argentinski paleontolog F. Ameghino je, doduše, opisao više, tobože tercijarnih ostataka čovjeka iz Južne Amerike. Radi se o komadima lubanja, komadima nekih drugih kosti, dalje o prvom vratnom kralješku (atlasu), nađenom na Monte Hermoso i dr. Ameghino je na tim ostatcima, koje pripisuje ljudskim predšasnicima, nazvanima Tetraprothomo, Triprothomo, Diprothomo i t. d., osnovao tvrdnju, da se čovjek u toku tercijarnog doba razvio u Južnoj Americi. Kako je to napose Hrd1ička pokazao, svi ti ostatci potječu od sadašnjeg čovjeka, komad bedrenjače pak od velikog grabežljivca iz porodice mačaka, a starost je onih ležišta nesigurna. Radi se, dakle, o tome, da je inače tako zaslužni Ameghino s onim svojim tvrdnjama potpuno zastranio.
VI. NAZORI O PREDŠASNICIMA ČOVJEKA.
Na osnovi činjenica, koje pružaju paleoantropologija, anatomija i embriologija, izražena su o podrijetlu čovjeka kao fizičkog bića brojna mišljenja, koja možemo svrstati u dvije grupe: poligenistička (polifiletička) i monogenistička (monofiletička). Poligenisti smatraju, da je današnji ljudski rod produkat više konvergentnih loza, koje su se već u donjem tercijaru odcijepile od zajedničkog debla, i da je svaka od tih loza bila izvorom nekih majmuna i neke ljudske grupe.
Glavni je predstavnik poligenizma bio H. Klaatsch (1910), koji drži, da su se gorila i neandertalac razvili iz zajedničke loze, koja se dalje nastavila s Crncima, a druga, orangoidna loza, da je dala izvor orangu i preko aurignacske rase Mongolima. Sličnim nazorima naginje Th. Ar1dt (1915), koji zastupa hipotezu, da su se od eocenskih Tarsioidea odcijepile 3 grane, jedna, kojoj bi vršci bili Cebidi, Semnopithecini, orang i Mongoli, druga, koja vodi Hapalidama, gibonima, izumrlom Dryopithecus-u, čimpanzi, Pithecanthropus-u i europidnoj rasi, i treća, koja je dala izvor izumrlom Sivapithecus-u, gorili i Crncima. G. Sergi (1913) predviđa, dapače, pet razvojnih loza, o kojih je jedna Dryopithecus s Neandertalcem i heidelb. . čovjekom) izumrla, isto i druga (neki Ameghinov »pračovjek«), samo što je ostao od nje odvojak, iz kojeg su se razvili kao posebna loza američki urođenici. Treća loza obuhvaća gorilu, čimpanzu, Crnce i bijelu rasu, a četvrta orangutana i Mongole.
Poligenistički pokušaji nisu našli mnogo pristaša, jer je, kako je to napose H. V. Vallois iscrpljivo objasnio, isključeno, da bi mogle sve brojne prilagodbe, koje su tipično ljudske i zajedničke svim sadašnjim ljudskim rasama, u više navrata neovisno nastati. Golema većina stručnjaka zastupa stoga monogenističko gledište, naime, da je današnji ljudski rod jedinstvenog podrijetla, ma da se razilaze nazori o tome, u koje se je doba i iz koje grane zajedničkog debla primata odvojila ljudska loza.
Što se neposrednih predšasnika čovjeka tiče, to se pripisuje glavno značenje fosilnim ostatcima, ma da ima i u njihovu ocjenjivanju različitih mnijenja. Najviše pristaša ima nazor, da je čovjek prošao morfološke stupnjeve, koje nam predočuju Pithecanthropus, Sinanthropus, Homo neanderthalensis i Homo sapiens fossilis. Da Pithecanthropus spada u izravnu lozu čovjeka, za to se već izjavio E. Duboisis njime svi stručnjaci, koji u Pithecanthropus-u vide prelaznu formu; napose je pokušao Schwalbe (1899) točnom anatomskom analizom potkrijepiti nazor, da je razvitak čovjeka pošao preko Pithecanthropus-a i Neandertalca. Kasniji su nalazi taj nazor još upotpunili, pa ako upoređujemo profile lubanja, pruža nam se od Pithecanthropus-a do čovjeka gotovo potpuna razvojna serija. Napose i E. Fischer i H. Weinert zastupaju to mišljenje.
No time se ne postavlja tvrdnja, da bi bio baš onaj Pithecanthropus, čija nam je lubanja sačuvana, ili baš jedan od pronađenih Sinanthropus-a ili Neandertalaca naš neposredni predšasnik. Pa bio bi zbilja malo vjerojatan slučaj, da bi se baš ostatci onih individuuma sačuvali, koji sačinjavaju neposrednu ljudsku lozu. Nego one ostatke shvaćaju vrlo malo promijenjenim potomcima izravnih predšasnika čovjeka, isto tako kao što se današnji Australci smatraju malo promijenjenim potomcima predšasnika svih ostalih ljudskih rasa. Jedino kao morfološki stupanj možemo Australce nazvati pređima ostalih ljudskih rasa, ma da time nitko ne misli kazati, da je neki sada živući Australac otac nekog Evropljanina. Zato i starost fosilnih nalaza hominida nipošto nema onu važnost, kakvu bi joj često htjeli pripisivati. Po tom mnogo zastupanom nazoru ljudski bi se predšasnici vrlo slični Pithecanthropus-u dalje razvili u forme poput Sinanthropus-a, ovi bi se upotpunili u pravcu Steinheimca i još dalje u pravcu Neandertalca, a konačno bi iz Neandertalcu sličnih forma nastao pravi čovjek. Eoanthropus, ako zbilja spadaju čeljust i lubanja zajedno, bio bi jedan postrani izumrli ogranak; jednako bi bio Homo rhodesiensis krajni ogranak loze, koja je drugim smjerom pošla od Neandertalca.
Ima i nazora, da Neandertalac ni morfološki ne spada u neposrednu lozu čovjeka. Već je Sera držao, da je to posebna forma čovjeka, prilagođena na ledeno doba; zato tako robustna okosnica. Ali to se ne slaže sa činjenicom, da je Neandertalac živio već kroz čitavo toplo Riss-Würmsko interglacijalno doba, a da ima i pravi čovjek iz ledenog doba. Ističe se također, da postoji izvjesna poteškoća za mnijenje, da je Neandertalac izravan predšasnik sadašnjeg čovjeka, u činjenici, da ima ostataka pravog čovjeka starijih nego su ostatci Neandertalca. Tu dolazi napose u obzir lubanja iz Swanscombe-a, pogotovu, jer se ona priključuje na steinheimsku lubanju.
Kojim bi putem bio pak pošao razvitak od nižih stupnjeva do stupnja Pithecanthropus-a, za to ima prilično malo oslona u okaminama. Ostatci primata poznati su sve od donjeg eocena dalje. Dosele je poznato od fosilnih primata: 15 rodova Chyromioidea, svi iz eocena, 12 rodova Lemuroidea, od eocena do diluvija, 22 roda Tarsioidea, svi iz eocena, 1 rod širokonosaca, Homunculus iz miocena, 4 roda Cercopithecina iz miocena i pliocena, 3 roda Semnopithecina iz pliocena, 4 roda Hylobatina, od oligocena do pliocena i 7 rodova Anthropopithecina iz miocena i pliocena. Radi se većinom o zubima ili donjoj čeljusti, ima sačuvan i koji desetak lubanja i par okosnica, dakle vrlo skučen materijal. Jedan od najstarijih fosilnih primata je Tarsioid Tetonius (Anaptamorphus) iz donjeg eocena u Wyomingu, kojemu su već Cope, a kasnije i mnogi drugi, pripisivali naročito značenje kao zajedničkom predšasniku i majmuna i čovjeka. Za međusobne veze antropomorfa ima nešto više podataka. Za gibone drži Schlosser, da im je praotac Propliopithecus iz oligocena, na kojega povezuje Pliopithecusa iz miocena, a dalje sadašnje gibone. Više ostataka antropomorfa, zubi i donjih čeljusti, iz miocena u Evropi i Indiji pribraja se rodu Dryopithecus. D. Fontani iz Francuske pokazuje izvjesne sličnosti sa zubalom gorile, D. germanicus ima više sličnosti sa čimpanzom, a kutnjaci D. Darwini naliče na čovječje, tako da i Gregory i O. Abel i H. Weinert dolaze do zaključka, da njegov položaj nije daleko od loze hominida. S Dryopithecus-om je već Dubois doveo u srodstvenu vezu Palaeopithecus-a iz donjeg pliocena u Indiji, od kojega je sačuvana gornja čeljust; Gregory pak smatra, da je Palaeopithecus forma bliska gorili. Drugi veliki antropomorf, Sivapithecus, poznat je iz miocena u Indiji, i to po komadima čeljusti. Kutnjaci su po svom obliku sličniji čovječjima nego kutnjaci sadašnjih antropomorfa, tako da je G. E. Pi1grim (1915) pokušao ovu formu smjestiti među hominide, što se međutim ne slaže s oblikom očnjaka, koji strši preko ostalog zubala kao kod ostalih antropomorfa. Kod Taungsa u Južnoj Africi, u ležištu iz donjeg diluvija, pronađena je 1924 dječja lubanja uz dio donje čeljusti. R. A. Dart, koji ju je ispitao, nazvao je taj nalaz Australopithecus africanus. Sadržinu lubanje odredio je Darts 520 cm3, što bi odgovaralo sadržini odrasle lubanje od 625 cm3, pa je bio stoga mnijenja, da je to neka prelazna forma k hominidima. Kasnije je W. Abel odredio sadržinu s 390 cm3, odrasla lubanja imala bi prema tome tek 450 cm3; naročito s obzirom na zubalo W. Abel dolazi do zaključka, da Australopithecus ne stoji u neposrednoj vezi s lozom hominida. G. 1937 pronašao je R. Broom kod Sterkfonteina u Transvalu ležište iz srednjeg diluvija različne ostatke slične, ali gotovo odrasle forme. Najvažniji je komad gornje čeljusti, koji pokazuje, da je zubalo bilo izvanredno slično čovječjemu, jer očnjak ništa ne iskače iznad ostalih zubi. S obzirom na zubalo Broom je za tu formu predložio naziv Plesianthropus. Drugu ovoj blisku formu pronašao je Broom 1938 kod Kromdraaia nedaleko od Sterkfonteina. Radi se o većem dijelu lubanje, koje se sadržina procjenjuje sa 600 cm2. Lubanja pripada velikom antropomorfu, ali je zubalo opet nalik na čovječje. Iz oblika zatiljnog zgloba proizlazi, da je ta forma hodala već poluuspravno. Taj je nalaz nazvan Paranthropus robustus. Ležište je iz gornjeg diluvija. Sve to ukazuje na to, da se radi o posebnoj izumrloj lozi antropomorfa, koja je morfološki bliža čovjeku nego sadašnji antropomorfi.
Kako bi imali zamišljati srodstvene veze čovjeka s ostalim primatima, u zadnje su vrijeme pokušali izraziti rodoslovnim crtežima H. F. Osborn, W. K. Gregory, A. Naef i H. Weinert.
Osborn drži, da se čovječja loza u oligocenu odijelila od loze, koja vodi k antropomorfima, i to s obzirom na kameno oruđe iz Red-craga i Fox-halla u Engleskoj. Moir, koji je pronašao to već dosta savršeno oruđe, drži, da potječe iz pliocena, iz česa bi proizlazilo, da su se hominidi razvili već mnogo ranije. Loze hominida i antropomorfida bile bi po nazoru Osborna dvije usporedne grane, kakve nam i inače paleontologija često predočuje kod različnih drugih grupa sisavaca. Napose upozoruje i W. E. Le Gros C1ark na to, da treba pretpostaviti kod primata više usporednih razvojnih loza, pa da međusobne sličnosti ne moraju biti izraz neposrednog srodstva, nego samo jednako ili slično usmjerenog usporednog razvitka.
Gregory drži, da je loza hominida i velikih antropomorfa bila sve do miocena zajednička, dok bi se loza gibona već na početku oligocena bila ocijepila. I Naef stavlja razvitak hominida u tercijarno doba, i to na svršetak miocena, te drži, da se loza antropomorfa odvojila od ljudske već sredinom oligocena, ali po njegovu nazoru dolazi poslije odvajanja antropomorfa još jedan nepoznati razvojni stupanj na početku miocena, koji u daljnem razvitku napokon dovodi do stupnja hominida.
Weinert, čiji su crtež rodoslovlja primata prihvatili i mnogi drugi antropolozi (E. Fischer, Verschuer, Eickstedt i dr.), stavlja odvajanje loza hominida antropomorfa u mnogo kasnije doba, u pliocen. On napose ističe kako od svih antropomorfa ima čimpanza najviše zajedničkih obilježja s čovjekom: poredak izlaska žila kucavica iz luka glavne odvodnice (aorte) jednak je kao kod čovjeka, što vrijedi, doduše, i za gorilu; šupljina u čeonoj kosti ima oblik, iz kojega se može izvesti oblik te šupljine kod Pithecanthropus-a i dalje kod Neandertalca i sadašnjeg čovjeka; poseban otvor, foramen spinosum, na donjoj strani lubanje za prolazak nekog živčića i nutarnje čeljusne kucavice postoji kod čimpanze jednako često kao kod čovjeka, dok je kod gorile rjeđi; mjesto, gdje se na lubanji pričvršćuju mišići šije, kod čimpanze je izbočeno kao kod čovjeka, dok je kod gorile i oranga plosnato; bubreg čimpanze ima isto kao čovječji više papila, dok gorila i orang imaju samo 1 papilu. Trudnoća traje kod čimpanze isto kao kod čovjeka, 280 dana. Naročita se važnost pripisuje Uhlenhuth-Nuttalovoj reakciji, iz koje slijedi, da je krvni serum čimpanze gotovo istovjetan s čovječjim, dok gorila i orang pokazuju jasne razlike; ako pomiješamo s antiserumom za čovjeka serume čovjeka i različnih antropoida, odnošaj je u količini taloga, izražen u brojkama: za serum čovjeka i čimpanze 10—12, za serum gorile 6, oranga 4, za pavijana 4—3. U tom pogledu se navodi, da se brojevi izomera jedne te iste bjelančevine kreću u kvadriljonima i kvintiljonima, pa izgleda gotovo nemoguće (vjerojatnost tek kojih 10-24), da bi se mogao javiti tu i tamo isti izomer neovisno, uslijed neke konvergencije ili sl. Do sličnih zaključaka kao Weinert dolazi i A. Keith. Keith je cijeli niz anatomskih obilježja izrazio točkama, pa je našao, da čovjek ima sa čimpanzom 396 točaka zajedničkih, s gorilom 385, s orangom 272, a s gibonom tek 188. Iz tih bi se činjenica moglo zaključiti, da postoji između čovjeka i čimpanze bliža filogenetska veza nego s ostalim antropomorfima, da bi se, dakle, predšasnici čimpanze bili najkasnije odcijepili od loze hominida, i to po Weinertovu nazoru u gornjem pliocenu. Svakako, Osborn i Gregory griješe, kada stavljaju u svojim rodoslovnim crtežima gorilu bliže čovjeku nego čimpanzu.
Malo je pristaša našlo shvaćanje, koje je zastupao već Cope (1882), a zatim Hubrecht (1897), da bi se loza hominida već na početku razvitka primata bila odvojila od ostalih loza. Kasnije se za taj nazor zalagao Wood-Jones (1919). Potonji tvrdi, da se čovječja loza već u eocenu odcijepila od zajedničke loze, i to izravno od Tarsioida (makija), što motivira time, da maki hoda donekle uspravno stražnjim nogama. Ali taj uspravan hod kod makija prilagodba je na skakanje, kao kod klokana ili kod skočimiša, pa se ne može porediti s čovječjim hodom. Građa makijeve noge s produljenom petnicom i čunicom (calcaneus i naviculare) pokazuje, da je to specijalizirana grupa, koja nikako ne bi mogla spadati u neposrednu lozu čovjeka. Izravni razvitak čovjeka iz primitivnih primata zastupa i Montandon (1928), koji drži, da čovječja loza izravno potječe od Lemuroidea.
Sasvim izoliran je Westenhöfer (1930) sa svojim nazorom, da se ljudska loza već najranije odcijepila izravno od općeg tipa sisavaca. On to opravdava građom našeg stopala, koje da pokazuje, da su već naši najraniji predšasnici hodali po zemlji, a da su se forme, koje žive na stablima, kasnije odcijepile od te loze i promijenile, tako da bi se majmuni i polumajmuni razvili iz ljudskih predšasnika, a ne obrnuto. Još dalje ide E. Dacqué, koji, napose, u popularnim spisima, brani tezu, da su predšasnici čovjeka već od paleozoičkog doba tvorili zasebnu lozu, koja nema nikakvih veza s razvojnim lozama životinja. Ti predšasnici da su imali, doduše, u početku oblik sličan vodenjaku, kasnije sličan gmazu, ali da su imali ipak ljudska svojstva. On motivira taj svoj nazor pričama o zmajevima i aždajama, koje da su uspomene iz mezozoičkog doba, kada da su naši predšasnici živjeli među tadašnjim orijaškim gmazovima. To je, naravno, samo neka fikcija, jer iz onih doba nema nikakvih tragova kakvoga inteligentnog bića, pa taj nazor znanost uopće nije uvažila.
ČIMBENICI POSTANKA ČOVJEKA.
Kako većina stručnjaka drži, da su predšasnici čovjeka bili arborikolna bića, koja su provodila život na stablima, svi se nazori o čimbenicima ljudskog razvitka slažu u tome, da je bio jedan od uvjeta, koji su pospješili humanizaciju, taj, da su predšasnici čovjeka radi promijenjenih prilika bili prinuždeni ostaviti šume i provoditi život u otvorenim predjelima. Barrel i Osborn pripisuju važan utjecaj dizanju gorskog nabora zemljine kore, koji je stvorio današnje gorske lance od Alpa preko Kavkaza, Irana, Himalaje do kineskih gorja. Time su nastali sjeverno od tog gorja hladni i otvoreni predjeli, s mnogo većim zahtjevima na životnu borbu, što je prisililo čovječje pređe da promijene svoj život i prihvate uspravan hod. Hod na stražnjim ekstremitetima povlači za sobom njihovo produljenje (klokan, skočimiš, maki i t. d.) uz izvjesne modifikacije u građi. Zbog uspravnog hoda postala je ruka potpuno slobodna, pa se tako usavršila njena upotreba. Uspravan hod rasterećuje mišiće i veze šije, jer je lubanja u svom težištu poduprta kralježnicom. Kako drži Anthony, to bi bio jedan od momenata, koji su omogućili, da se lubanja proširi i mozak poveća. Da bi se mozak povećao tek poslije postignuća uspravnog hoda, potvrđuju, kako ističe Schwalbe, prilike kod Pithecanthropus-a, koji je imao već uspravan hod, ali tek mali opseg lubanje. Bolk pripisuje povećanje mozga t. zv. fetalizaciji (fetus »zametak«). On polazi s gledišta, da čovjek u pogledu obujma mozga više sliči embrionima antropomorfa nego odraslim formama, pa stoga drži, da je bitan činilac kod stvaranja čovjeka bila pojava, da su njegovi predšasnici zadržali izvjesna embrionalna obilježja, napose s obzirom na mozak. Uz to Bolk pripisuje naročitu ulogu promijenjenom djelovanju endokrinih žlijezda, pogotovu onih, koje utječu na rast ili pospješuju razvitak (hipofiza i štitnjača). Bolk drži, da je taj momenat doveo do toga, da su se stanice u mozgu još jedamput razdijelile i tako svoj broj podvostručile. Povećanje bi mozga, dakle, uslijedilo jednim korakom. Promijenjenom djelovanju endokrinih žlijezda pripisuje Bolk i gubitak dlaka na tijelu. Za to, da su promjene u endokrinom djelovanju imale važnog utjecaja, zalaže se i Versluys. Protiv hipoteze o fetalizaciji oštro je istupio Neuville, koji ističe, da je opća pojava u embrionalnom razvitku svih životinja, da se centralno živčevlje ranije osniva nego ostali organi, pa svi embrioni imaju relativno veći mozak nego odrasli stadiji. Weinert nalazi u otkriću pravljenja vatre momenat, koji da je pospješio razvitak čovjeka i omogućio njegovu sve veću nezavisnost od klimatskih prilika, a uživanje vatrom priređene hrane da je povuklo za sobom umanjenje zubala, napose očnjaka, i mišića za žvakanje, što se s obzirom na zubalo ne slaže s time, da imamo i kod Australopithecus-a već izravnano zubalo i smanjene očnjake. Većina vidi važan poticaj za usavršavanje čovjeka u teškim klimatskim prilikama ledenoga doba, koje su iziskivale sve veće sposobnosti u borbi s prirodom.
MJESTO POSTANKA ČOVJEKA.
Za pitanje, gdje je bila kolijevka ljudskoga roda, mjerodavno je, da su hominidi bili već u srednjem diluviju zastupani i u Europi i istočnoj Aziji (Heidelberg, Peking), pa se stoga zaključuje, da je mjesto postanka čovjeka imalo središnji položaj u starom svijetu, tako da su se hominidi mogli odatle proširiti na sve strane. Uz to se pretpostavlja, da su to bili hladni predjeli bez šuma. Tako većina stručnjaka drži, da treba kolijevku hominida tražiti u središnjoj Aziji, negdje sjeverno od Himalaje. Weinert pak više naginje shvaćanju, da bi mogla biti kolijevka hominida u Evropi, i to s obzirom na brojne stare nalaze u Europi, ne uvažuje pak kod toga, da su tako brojni ostatci hominida pronađeni u Evropi i s razloga, što je evropsko tlo najviše prekopavano i istraživano. Montandon, koji stoji na gledištu hologeneze, kako je naučava Rosa, dosljedno tom svom shvaćanju drži, da uopće nema određenog mjesta stvaranja čovjeka, nego da su se po načelima hologeneze na svim kontinentima u određeno doba iz lemuridima sličnih predšasnika čovjeka stvarali hominidi, ali u tom svom shvaćanju nema mnogo sljedbenika. Nekakvoj hologenezi, ali u mnogo manjem opsegu, naginje u zadnje vrijeme F. Weidenreich. On drži, doduše, da su hominidi jedinstvenog, monofiletičkog porijetla, ali zastupa mnijenje, da se čovjek dalje razvijao policentrički, naime na više mjesta, tako Cromagnonac u Maloj Aziji (s obzirom na nalaze u Palestini), Crnci u Africi kao potomci Rodezijca, Mongoli u Kini iz Sinanthropus-a, a Australci na Sundskim otocima iz javanskog pračovjeka.
LIT.: O. Abel, Die Stellung des Menschen im Rahmen der Wirbeltiere, Jena 1931; L. Bolk, Das Problem der Menschwerdung, Jena 1926; M. Boule, Les hommes fossiles, Pariz 1923; Ch. Darwin, The Descent of Man and on Selection in Relation to Sex, London 1871 (njemački prijevod: Die Abstammung des Menschen, Leipzig, Reclam); K. Gorjanović-Kramberger, Der diluviale Mensch von Krapina, Wiesbaden 1906; Human Biology and Raciale Welfare, Ed. by E. V. Cowdry, New-York 1930; H. Klaatsch, Der Werdegang der Menschheit, Berlin 1920; L. S. B. Leakey, Stone Age Africa, London 1936 (njemački prijevod: Steinzeit-Afrika, Stuttgart 1938); A. Naef, Die Vorstufen der Menschwerdung, Jena 1933; G. Schwalbe, Die Abstammung des Menschen und die ältesten Menschenformen; Die Kultur der Gegenwart III/5, Leipzig 1923; B. Škerlj, Človek, Ljubljana 1932; H. Weinert, Ursprung der Menschheit, Stuttgart 1932; Isti, Entstehung der Menschrassen, Stuttgart 1938; E. Werth, Der fossile Mensch, Berlin 1928; R. Wiedersheim, Der Bau des Menschen als Zeugnis für seine Vergangenheit, Tübingen 1908.
D. Čovječje rase.
I. POJAM I DEFINICIJA RASE.
Rasa je skupina pripadnika iste prirodne vrste, koji se, osim u obilježjima svoje vrste, podudaraju još u izvjesnom broju daljnjih nasljednih osebina, te se po tim osebinama razlikuju od drugih sličnih skupina. Nasljedna svojstva su, dakle, mjerodavna za rasu, a nipošto ne raznolike njihove variacije proizvedene utjecajem okoline. Već se i prije naglašavalo značenje nasljednih svojstava za pojam rase. O njima govore već Kant (1785), Prichard (1839), De Quatrefages (1867), Topinard (1885), dok su drugi naglašavali zajednička somatička svojstva uopće, ili zajedničko podrijetlo. Neki su, tako već G. R. Forster (1786), rasu jednostavno identificirali s narodom, a napose su se kod manjih rasnih grupa pridržavali lingvističke podjele. Uvid u pravu bit rase, kako je izražena gornjom definicijom, omogućen je tek u ovom stoljeću napretkom nauke o nasljeđivanju, koja je objasnila, da su nasljedstveni faktori ono, što sačinjava suštinu organizma, dok su somatička svojstva tek jedan odraz tih faktora. Somatička su obilježja samo neki indikator za nosioce nasljedstva, jer ona ovise i o utjecaju vanjskih čimbenika, okolini, u kojoj se organizam slučajno nalazi. Opažati možemo, doduše, samo somatičke osebine, pa je, dakle, za ispravno razlikovanje rasa od važnosti, da se iz somatičkih svojstava izluči ono, što je proizvedeno utjecajem okoline, kako se ne bi u rasne značajke uvukle i modifikacije uzrokovane vanjskim čimbenicima.
II. POSTANAK RASA.
O stvaranju rasa u živoj prirodi donijela je brojnih iskustava eksperimentalna genetika Morganova smjera u vezi s genetičko usmjerenim biogeografskim istraživanjima, na kojima mnogo rade i ruski genetičari (T. Dobžanski, Dubinin, Romašov i dr.), a kod nas Z. Lorković. I matematički je taj problem obrađen u tančine (S. Wright, J. B. S. Haldane, R. A. Fisher).
Prvobitno jedinstvena skupina pripadnika neke vrste cijepa se na rase uslijed mutacija, t. j. promjena nasljedstvenih čimbenika, koje se stalno spontano javljaju kod pojedinaca. Ako dolazi do izolacije manjih skupina, to može odrediti već puki slučaj, da daljnjim križanjem mutiranih pojedinaca s nepromijenjenima uzmu sve više maha nova svojstva. Napose pak formirat će se nova rasa, ako vanjski čimbenici na nekom području pogoduju nosiocima jedne vrste promijenjenih svojstava. Što je brojnija neka vrsta (species), što je većim područjem proširena i što ima više raznolikih uvjeta za život na tom području, to će biti više mogućnosti za stvaranje rasa. Pogotovo će se stvarati nove rase, ako dođe do seljenja većih skupina tako, da postignu nove životne uvjete, koji će više pogodovati drugim mutacijama, nego životni uvjeti u starom području. Slično će djelovati promjena životnih uvjeta u istom području radi mijenjanja klime, kao što je to bilo u ledenim i međuledenim dobima. Od velikog su značenja i geološke promjene zemaljske površine, dizanje bregova i transgresije mora, koja rastavljaju prije spojene zemlje.
To zbivanje, da se javljaju nove mutacije i time nove mogućnosti za bolje prilagođivanje okolini, nastavlja se stalno dalje. Rase, dakle, nisu ništa postojano, nego su, tako reći samo faze u tom zbivanju i podvrgnute su sve daljem cijepanju i preobražavanju, naravno, tek u razdobljima od više tisuća generacija.
Što su rase starije po svom postanku, to će veće i jasnije biti razlike između njih. Mlađe će skupine imati više zajedničkih svojstava, a bit će u svakoj skupini još velik postotak vrlo sličnih pojedinaca. Zbivanje, koje dovodi do stvaranja rasa, ne zahvaća, naime, odmah sve pojedince neke skupine, nego tek mali broj, koji će se s vremenom, zbog djelovanja prikazanih čimbenika, povećavati. Mada nije još cijela skupina jedinstvena, mogu djelovati već daljnji izolacioni i migracioni procesi, i nastupit će novo cijepanje, ma da se prvo još nije dovršilo. Do »čistih« rasa doći će stoga vrlo rijetko, jer će te skupine stalno još sadržavati neki postotak pojedinaca, koji će pokazivati osebine prvobitne skupine, a prije nego svi takvi iščeznu, javit će se već nova cijepanja.
Za stvaranje novih rasa važno je i križanje onih, koje već postoje. Kod toga dolazi do novih raznolikih kombinacija među nasljednim osebinama izlaznih rasa, a za te kombinacije vrijedi isto, što za mutacije, da će se naći među njima takve, kojima će pogodovati uvjeti okoline, pa će se sve dalje množiti. Baš među čovječjim rasama ima nekih, koje su, bez sumnje, proizvod križanja dviju vrlo raznolikih rasa, kao što su na pr. Etiopidi, Turanidi i različne skupine u Istočnoj Indiji.
Izloženi uvjeti, koji su mjerodavni za sve organizme u slobodnoj prirodi, znatno se mijenjaju, ako je neka skupina zaštićena od različnih nepovoljnih utjecaja prirode, kao što su domaće životinje, koje čovjek njeguje i čuva. Svakojaka svojstva, koja se ne mogu održati u divljem stanju, jer se tu njihovi nosioci gotovo istrijebe, moći će se sačuvati pod zaštitom čovjeka, na pr. najraznovrsnije boje dlaka. Volja čovjekova odlučuje kod domaćih životinja, koje će se kombinacije osebina i koja imitirana svojstva održati, jer čovjek izabire za rasplod nosioce onakvih nasljednih svojstava, koja smatra prikladnima za svoje potrebe. Zato nalazimo kod domaćih životinja kud i kamo veću pocijepanost na rase nego kod sličnih životinja u divljem stanju. U donekle sličnom položaju kao domaće životinje nalazi se i ljudski rod, na što je napose upozorio E. Fischer (1914). Čovjek je, naime, već od davnine u stanju neke autodomestikacije, jer udruživanje pojedinaca u skupove, u kojima si pružaju međusobnu pomoć, omogućuje, da se dalje rasplođuju nosioci onakvih mutiranih svojstava, koja se ne bi mogla sačuvati u slobodnoj prirodi izvan ljudskog društva.
Zaštita, koju je pružao socijalni život već od prvih njegovih početaka, svakako je, dakle, uvelike ublažila djelovanje nekih momenata, koji surađuju kod stvaranja rasa. Unutar skupina, koje su bile izvorom novih rasa, moglo se je stoga održati mnogo više pojedinaca s osebinama izlazne populacije, tako da, pogotovo kod čovjeka, nikad nije moglo doći do stvaranja »čistih« rasa, naime skupina, gdje bi svi pojedinci bili opremljeni osebinama, koje su mjerodavne za pravac razvitka.
Procesi stvaranja čovječjih rasa mogli bi se prema izloženome zbivati samo dotle, dok je bila zemlja još rijetko naseljena, dok je mogla djelovati borba s prirodom i međusobna borba plemena u smislu očuvanja nosilaca podesnih svojstava. Kada je međusobna zaštita postigla već toliku mjeru, da su se formirala veća plemena i napokon narodi, morali bi prestati gotovo svi procesi stvaranja novih rasa na osnovu prednosti, koje pružaju izvjesne mutacije pojedinaca. Ostao bi samo momenat, koji istakosmo već na početku, a to je izolacija manjih skupina. Kako je pokazao Wright matematičkim izvodima, u takvim skupinama puki slučaj mora dovesti do toga, da u jednoj skupini s vremenom prevlada jedno novo svojstvo, a u drugoj skupini drugo, pa tako dolazi bez sudjelovanja kojih drugih utjecaja do razilaženja pojedinih skupina u njihovim obilježjima. Ove poteškoće pokušava uz to prebroditi hipoteza ortogenetskog razvitka, kako je zamišlja i E. Fischer. Mutacije, promjene nasljednih faktora, uzrokovane su još gotovo nepoznatim izvanjim utjecajima, koji djeluju svakako neposredno na nasljedne faktore, da ih promijene. Svaka bi okolina izazivala ponajviše svojevrsne mutacije, pa bi stoga boravak u istoj okolini stalno proizvodio u istom pravcu usmjerene mutacije, tako da bi se započeti razvitak automatski nastavljao u dotičnom pravcu. Tako bi unatoč zaštiti nemutiranih pojedinaca mogao, kroz dugi niz generacija, oteti maha neki novi nasljedni tip, jer bi mutacije zahvaćale sve više i više pojedinaca.
Svako je obilježje proizvod nasljedstva i okoline. Napredak civilizacije i kulture stvara stalno nove životne uvjete, što se mora odraziti i na somatičkim osobinama. U slijedećem poglavlju upozorit ćemo na neke vrlo značajne rezultate u tom pogledu.
U gornjim izvodima prikazani postanak rasa osniva se na iskustvima postignutim eksperimentalnim proučavanjem mutacionih pojava, kao i proučavanjem mutacija u prirodi na različnim podesnim životinjama i biljkama, dakle proučavanjem mikroevolucije, t. j. djelovanja onih malih nasljednih promjena, koje se stalno javljaju kod organizama, te su pristupačne eksperimentu. Većina biologa, među njima i većina u pravcu nauke o nasljeđivanju usmjerenih antropologa, shvaća razvitak rasa i vrsta, kao uopće i cijelo evoluciono zbivanje, u smislu tih eksperimentalnih iskustava.
Mnogo paleontologa, koji imaju pred očima velike promjene organizama u toku geoloških perioda, drže, da postoji i neposredno prilagođivanje, koje s vremenom biva nasljedno. Za ovo shvaćanje zalaže se i Eickstedt. Po tom mišljenju od važnosti su kod stvaranja rasa pored ostalih evolucionih mehanizama i promjene, koje nastaju uslijed utjecaja izvanjih čimbenika. Ako, naime, ovi djeluju kroz mnogo generacija stalno u istom smislu, izazvane bi promjene postale nasljedne. Priznati treba, da bi ovakvo zbivanje mnogo jednostavnije i brže dovelo do stvaranja novih jedinstvenih rasa nego lih kumulacija mikroevolucionih zbivanja. Onda otpadaju sve poteškoće i zapreke, koje proizlaze za stvaranje rasa iz socijalnih momenata. Tako bismo mogli vrlo jednostavno razumjeti pojačano stvaranje pigmenta, kakvo se očituje kod Crnaca i Melanezida, naime, da je stalno obasjavanje tropskim suncem dovađalo kod svakog pojedinca tokom njegova života do umnažanja pigmenta, a da je ta na koži izazvana promjena djelovala nekim još nepoznatim čimbenicima na nasljedstvenu tvar te je s vremenom promijenila u smislu, da je postalo svojstvo jače pigmentacije nasljedno.
Ali svi dosele provedeni pokusi, napose pokusi s mušicom Drosophila, koja ima na godinu do 20 generacija, što odgovara kod čovjeka razdoblju od 600 godina, nisu donijeli nikakvog rezultata, koji bi govorio u prilog takvom tumačenju; svi pokusi, koji bi mogli poslužiti kao dokaz za nasljeđivanje svojstava postignuti utjecajem okoline, pokazali su se kod točnijeg ispitivanja nedostatnima. Eickstedt i drugi pristaše ovog nazora upozoruju na vremenski čimbenik (faktor), koji da nije dostatno uvažen, jer da treba kud i kamo dužih razdoblja od više tisuća generacija, da bi svojstva, inducirana od okoline, postala nasljedna.
Kao daljnji čimbenik kod stvaranja rasa navodi Eickstedt »harmonizaciju« osebina. On pretpostavlja neki posebni mehanizam kod djelovanja nasljednih faktora, koji da dovodi do toga, da se kod križanja rasa time kombinirani nasljedni faktori obadviju rasa među sobom tako uravnoteže, da se generacijama bez naročitog izbora i eliminacije harmoniziraju svojstva križanaca, usklade u novu harmoničku cjelinu, koja će tvoriti novu rasu. On upozoruje u tom pogledu na Etiopide, koji su očito proizvod križanja bijele i crnačke rase.
Montandon, koji stoji na gledištu hologenetskog razvitka, tumači stvaranje rasa pod vidikom nutarnjih čimbenika, koji automatski, u smislu hologeneze, djeluju u izvjesnom pravcu i dovode prema izvanjim uvjetima do stvaranja novih nasljednih svojstava i time novih rasa. I tu bi igrao vidnu ulogu vremenski faktor, koji nije pristupačan eksperimentu.
Čovjek se već prije više desetaka tisuća godina proširio svim kopnima od ekvatora do njihova krajnjeg juga i daleko preko sjevernog polarnog kruga, u svim raznovrsnim klimama s brojnim raznolikim uvjetima života. Visoki lanci bregova i široki morski putovi ili čitavi oceani dijele ta obitavališta čovjeka, te su još donedavno činili gotovo neprolazne zapreke između pojedinih područja. Uz to znamo, da su se već u pretpovijesna vremena zbivale seobe čitavih populacija. Razumljivo je stoga, da kod čovjeka nalazimo s obzirom na nasljedna svojstva izvanredno veliku mnogoličnost, tako da nijedan čovjek nije jednak drugome i da prema glavnim nasljednim somatičkim obilježjima možemo svrstati ljudski rod u velik niz rasa.
III. NASLJEDNE RAZLIKE ČOVJEKA
Kako već istakosmo, kao kod svakog živog bića, i kod čovjeka fizička su svojstva proizvod dvaju čimbenika: nasljedstva i okoline. Kod svakog obilježja surađuju ova dva čimbenika, ali su neka obilježja više podvrgnuta utjecaju okoline nego druga. Što je manji utjecaj okoline, to su takva obilježja prikladnija za razlikovanje rasa.
Kao rasnom obilježju, veliko značenje pridaje se i visini tijela, koja je u biti, kao većina onih obilježja, koja mjerimo, vidljivi izražaj građe kostura. Svakako, nasljeđivanje ima veliku važnost za visinu tijela. Ali treba imati na pameti, da je kostur u svom rastu ovisan o različnim čimbenicima, koji su podvrgnuti vanjskim utjecajima. Na rast utječe prehrana, a utječu i endokrine žlijezde, napose hipofiza, koja u svom djelovanju opet ovisi o drugim žlijezdama s nutarnjim lučenjem, a na sve te sustave djeluju i vanjski čimbenici. Kako je Bo1k ustanovio, Holanđijci su zadnjih 70 godina postali veći prosječno za 10,9 cm. Valjda su tome bile uzrokom poboljšane ekonomske prilike. Irisava je također našao, da su japanski novaci sada prosječno veći nego prije 30 godina. Kod Indijanaca u Sjev. Americi, koji su preseljeni iz brda u nizinu, pokazalo se, da su postali veći, nego što su njihovi suplemenjaci, koji su ostali u brdima. U većini kulturnih zemalja pokazalo se, da su stanovnici gradova veći nego narod na selu. Visina tijela ima, svakako, kod istog nasljedstva varijacionu širinu od kojih 15%.
Kod normalnih ljudi različnih rasa visina se kreće između 121 do 209 cm. Kod većine rasa žena je za 6—8% niža od muškarca. Slijedeća tabela prikazuje poprečnu visinu (u cm) nekih populacija (po raznim autorima):
| Etnička skupina |
muš. |
žen. |
Etnička skupina |
muš. |
žen. |
| Mavambo-Pigmeji (Afr.) |
140,8 |
135,6 |
Sjev. Kinezi |
167,6 |
— |
| Negriti (Ist. Indija) |
146,3 |
137,8 |
Poljaci |
167,9 |
— |
| Senoi (Malaka) |
152,0 |
142,0 |
Maori (Nova Zel.) |
168,0 |
— |
| Laponci (Norv.) |
152,3 |
145,0 |
Slovenci |
168,3 |
— |
| Veda (Cejlon) |
153,3 |
143,3 |
Armenci |
168,5 |
152,9 |
| Ainu (Sahalin) |
156,7 |
147,1 |
Apaši (Amerika) |
169,1 |
156,8 |
| Polarni Eskimi |
157,0 |
145,0 |
Hrvati sa sjev. i istoka |
169,5 |
— |
| Japanci |
158,5 |
145,0 |
Ukrajinci . |
170,1 |
— |
| Siamci |
159,9 |
— |
Hrvati iz Dalmacije |
170,8 |
— |
| Južni Kinezi |
162,2 |
149,8 |
Šveđani |
170,9 |
— |
| Tatari s Volge |
162,8 |
— |
Srbi iz Srbije |
170,9 |
— |
| Talijani |
164,0 |
— |
Norvežani |
172,1 |
— |
| Francuzi |
164,1 |
157,0 |
Sioux-Indijanci |
172,6 |
159,5 |
| Cigani iz Rumunjske |
164,9 |
153,2 |
Englezi |
172,8 |
— |
| Bavarci |
165,6 |
— |
Hrvati iz Hercegovine |
173,3 |
— |
| Eskimi s Alaske |
165,8 |
155,1 |
Škoti |
174,6 |
— |
| Velikorusi (Tver) |
166,0 |
154,0 |
Masai-Crnci |
174,7 |
|
| Crnci iz Toga |
166,1 |
— |
Patagonci |
175,0 |
— |
| Bugari |
166,5 |
155,7 |
Malajci iz Sumatre |
175,5 |
— |
| Hrvati iz Istre |
166,8 |
— |
Crnogorci |
178,3 |
— |
| Nijemci iz Salzburga |
167,0 |
— |
Sara-Crnci |
181,7 |
167,6 |
Rasnim obilježjima smatraju se i različne proporcije tijela, omjer između širine ramena i visine tijela, između duljine trupa i udova, između duljine ruku i nogu, između pojedinih dijelova ruke i noge i dr. Sve ovakve proporcije izrazuju se u t. zv. indeksima, t.j. kvocijentom formulakraća mjera × 100/dulja mjera Indeks nam, dakle, izražava kraću mjeru u postotcima dulje. Velika se pozornost posvećuje obliku glave, odnosno lubanje, koja može biti, gledana odozgo, ili duguljasto ovalna, okruglasta ili gotovo sasvim okrugla, što se izražava s indeksom duljine i širine, dakle brojem, koji odgovara širini izraženoj u postotcima dužine: formula širina ×100/dužina. Što je indeks veći, to je glava okruglija. Tu govorimo o dolihocefalnoj (indeks do 75), mesocefalnoj (71,1—80), brahicefalnoj (80,1—85), hiperbrahicefalnoj (85—90) i ultrabrahicefalnoj glavi (iznad 90). Indeks duljine i širine, mjeren na lubanji, manji je od prilike za 1,5—2 jedinice od indeksa mjerenog na živom čovjeku.
Slijedeća tabela prikazuje prosječni indeks glave nekih etničkih skupina (po različnim autorima).
| Australidi iz sred. Australije |
71,9 |
| Melanezidi s Loyalty-otoka |
72,5 |
| Stari Španjolci |
73,2 |
| ,, Germani |
73,7 |
| ,, Englezi |
74,7 |
| ,, Rusi |
74,9 |
| Arnauti |
76.9 |
| Englezi |
77,6 |
| Irci |
77,9 |
| Norvežani |
78,0 |
| Španjolci |
78,1 |
| Balkanski Cigani |
78,2 |
| Sicilijanci |
79,6 |
| Bugari |
79,7 |
| Belgijanci |
80,5 |
| Letonci |
80,5 |
| Danci |
80,7 |
| Bjelorusi |
81,5 |
| Poljski Židovi |
81,8 |
| Velikorusi |
82,4 |
| Poljaci |
82,6 |
| Rumunji |
82,9 |
| Ukrajinci |
83,4 |
| Nijemci iz Badena |
83,6 |
| Nijemci iz Saske |
84,0 |
| Slovenci |
84,3 |
| Talijani iz Lombardije |
84,5 |
| Hrvati-muslimani |
84,6 |
| Hrvati sa sjevera i istoka |
85,2 |
| Armenci |
85,7 |
| Hrvati-katolici iz Bosne |
86,8 |
| Hrvati iz Dalmacije |
87,0 |
| Laponci |
87,6 |
| Kirgizi |
89,4 |
| Kalifornijski Indijanci |
89,7 |
Druga mnogo upotrebljavana mjera glave je morfološki indeks lica, koji izražava visinu lica od donjeg ruba brade do gornjeg kraja nosa u postotcima najveće širine, dakle formula visina lica × 100/širina lica. Taj se indeks kreće između 70 i 100; što je veći, to je lice dulje i uže.
Evo nekoliko indeksa lica:
| Kirgizi |
79,0 |
Kinezi |
87,0 |
| Filipinci |
81,3 |
Tirolci |
87,1 |
| Kamčadali |
83,3 |
Čukči (Sibirija) |
88,0 |
| Suaheli-Crnci |
83,3 |
Bjelorusi |
88,0 |
| Tunguzi |
84,4 |
Tatari |
88,4 |
| Krećani |
85,5 |
Eskimi |
88,8 |
| Nijemci iz Badena |
85,8 |
Švicari iz Safientala |
93,3 |
Ima cijeli niz drugih mjera i omjera glave. Ove metode mjerenja uveo je švedski antropolog A. Retzius (1842), te su brzo svestrano prihvaćene za izračunavanje rasnih razlika. S vremenom su se te metode izrodile. Tako je koncem 19. st. Török došao do nekih 5000 mjera, koje se mogu izvesti na lubanji. U novije vrijeme ne pripisuje se više toliko značenje tim različnim mjerama, jer se ustanovilo, da mnogo ovise o vanjskim utjecajima te da se u novoj okolini mogu promijeniti već u prvoj generaciji.
Kako je Boas ispitivanjem naseljenika u Sjev. Americi pokazao, omjeri glave ovise mnogo o vanjskim čimbenicima. Kod dolihocefalnih južnih Talijana biva glava u Americi nešto okruglija, a kod brahicefalnih Židova iz Poljske nešto dulja i uža; opažene razlike iznose do 2 jedinice indeksa.
Značajno je obilježje kut, u kojem se sastaju gornji i donji zubi, koji mogu stajati u okomitoj ravnini ili pak u tupom, s tjemenom prema vani okrenutom kutu, dakle koso, pa govorimo o ortognatiji (ravnozubosti), i prognatiji (kosozubosti). Potonja može imati svoj uzrok samo u koso smještenom položaju alveola (udubina za zubne korijene) ili u kosom položaju cijele čeljusti.
Dok su svojstva, koja ispituje antropometrija, određena oblikom kostura, to ima brojnih obilježja i u mekanim dijelovima; mnogo tih osebina prilično je manje ovisno o vanjskim utjecajima nego mjere kostura. Oblik nosa vrlo je značajno obilježje, napose njegov profil i njegova širina. Za prikazivanje širine nosa upotrebljava se indeks visine i širine nosa, koji izražava širinu u postotcima visine formula (širina×100)/visina što je indeks manji, to je nos uži.
Evo nekoliko indeksa nosa različnih etničkih skupina:
| Armenci |
60,4 |
Ukrajinci |
71,6 |
| Sjeverni Francuzi |
62,0 |
Kitajci |
72,9 |
| Eskimi |
64,1 |
Kalmiki |
73,9 |
| Gruzini |
64,5 |
Anamiti |
76,8 |
| Škoti |
65,1 |
Malajci |
82,6 |
| Nijemci iz Badena |
65,7 |
Filipinci |
83,8 |
| Oseti |
66,8 |
Kambodžani |
86,0 |
| Francuzi uopće |
67,3 |
Polinezijci |
89,8 |
| Grci |
67,6 |
Suaheli |
92,1 |
| Flamani |
67,7 |
Duala-crnci |
95,4 |
| Bjelorusi |
67,9 |
Sanidi (Bušmani) |
102,5 |
| Bugari |
68,1 |
Ašanti-crnci |
107,5 |
| Velikorusi |
69,9 |
Australci |
107,6 |
| Tatari |
70,8 |
Tasmanci |
108,9 |
| Cigani |
70,8 |
|
|
Usne također ne ovise mnogo o vanjskim čimbenicima. Njihov se oblik kreće između sasvim uskih, gdje se jedva primjećuje nešto crvenila sluznice, do debelo izbubrenih, koje su značajne za Crnce. Napose iskaču debele usne, ako su popraćene od prognatije. Oblik i razmjerna veličina uha pokazuju dostatnu mnogoličnost. Najdulju ušku ima dinarska rasa. Poseban oblik uha s odmaknutim gornjim krajem i priraslom jagodicom nalazimo kod Sanida (Bušmana). Formacija vjeđa i otvor za oko imaju mnogo različnih tipova. Ili su široko otvoreni ili čine među sobom usku pukotinu, a uz to su prema vani zategnute, kako se to opaža kod različnih Mongolida. Ako vjeđe čine širok otvor, taj može biti u obliku sferičkog dvokuta, ili s oštrijim kutom vani, a tupljim unutra, tako da otvor ima oblik badema, kako se opaža kod Orijentalida. Za Mongole je značajna formacija gornje vjeđe, koja ima poprečan nabor, a taj prelazi prema nosu u polumjesečni nabor, koji pokriva nutarnji kut oka, t. zv. mongolski nabor. Kod Hotentota vjeđe imaju mnogo masti, pa su debele kao kobasice. Oko može ležati duboko u očnoj šupljini ili je izbuljeno, što ovisi o količini masti iza oka. Rasno svojstvo je i formacija i položaj ženskih grudi, koje mogu biti okruglaste, čunjaste ili u obliku šiljate vreće, kao kod nekih crnačkih plemena. Razvitak i jačina mišića unutar svih rasa individualno su različiti; ovisni su mnogo i o vježbanju te radu, ali ima tu i nasljednih momenata. Kao rasno obilježje dolaze u obzir naročito mišići na sjedalu i na golijeni. Dok su prvi kod Europida napose dobro razvijeni, to se kod drugih rasa manje ističu. Mišići na golijeni kod Europida naglašeni su, dok su kod nekih crnačkih i američkih rasa manjeg opsega, tako da je golijen kao uzak stup.
Makar su razvitak i obujam potkožnog masnog tkiva u velikoj mjeri ovisni o prehrani, to ima i tu nasljednih momenata, naročito s obzirom na mjesto, gdje se gomila pričuvna mast. Kod većine rasa sakuplja se mast po cijelom potkožnom tkivu, ali najjače na trbuhu. Kod Hotentota i Bušmana nalazi se pak glavno mjesto sakupljanja masti na sjedalu. To svojstvo zovemo steatopigija (στέαρ »mast«, πυγή »stražnjica«). Da su i neke rase fosilnog čovjeka pokazivale nagnuće gojaznosti, odavaju primitivni kipovi, koje nam je ostavio fosilni čovjek. Opću ugojenost pokazuje ženska statueta, nazvana Venera od Willendorfa, proizvod Cro-Magnonskog čovjeka. Statueta, nađena u Mentone-u uz ostatke Grimaldi-rase, očituje jako razvijenu steatopigiju i još neke značajke, koje su tipične za Hotentote, tako da bi se moglo pomišljati na neke veze sa sadašnjim južnoafričkim urođenicima.
Daljnja skupina nasljednih osebina odnosi se na kožu i njezine tvorbe, vlasi i dlake. Sama koža može biti napeta ili naginje naborima, što je uvjetovano kakvoćom usmine, kožnog vezivnog sloja. Oštre nabore u licu nalazimo kod Dinarida, a kod Koisanida (Hotentota i Bušmana) počinje se koža već u srednjim godinama po cijelom tijelu, a napose u licu, slagati u brojne fine nabore. Dlakavost tijela kod većeg je dijela ljudskih rasa slabo razvijena. Najjaču dlakavost, a s njom i najbolji razvitak terminalnih dlaka te brade i brkova kod muškarca, nalazimo kod Europida, i to kod Ainua na Sahalinu i otoku Jeso, ali i neke evropske rase ne zaostaju mnogo za njima kao ni Australci. Kao naročito važno rasno obilježje smatra se oblik vlasi. Presjek vlasi nije kod svih rasa jednak. Ako je presjek okrugao, tada vlas strši kao slama. Što je presjek ovalniji, to vlasi više naginju valovitom i kudravom poretku. Korijen je vlasi u koži obično ravan; ako je ukrivljen, tada je vlas već od prvog početka, kada se stvara, ukrivljena, pa se njezina stabljika počinje spiralno kovrčati, čim dopre nad površinu. Kod najgušćeg kovrčanja pojedine se skupine vlasi među sobom spletu u tvorbe poput pupoljaka; to je t. zv. fil-fil kosa, kakvu nalazimo kod nekih Crnaca, a napose kod Koisanida.
Kompleksije se zovu ona obilježja, koja ovise o količini i kakvoći pigmenta (boje): boja kože, boja kose i dlaka te boja šarenice. Boja kože ovisi o vlastitoj boji površnog roženog sloja, koji je žućkast ili gotovo bezbojan. Kako je taj sloj providan, to manje ili više prosijavaju žilice vezivnog sloja, pa ima takva koža ružičastu boju. Ako je koža manje prozirna i ako nije tako nježna, naginje boja više na blijedo-žućkasto. U stanicama na donjoj strani površnog sloja može se stvoriti smeđi pigment u obliku malih zrnaca. Pigment se može stvarati i u vezivnom sloju, i to u naročitim pigmentnim stanicama. Prema količini i poretku pigmenta, prema prosojnosti površnog sloja i razvitku žilica u vezivnom sloju, koža pokazuje najraznovrsnije boje: žućkastu, svijetlosmeđu, crvenkasto-smeđu, tamnosmeđu poput kave, tamnosivkastosmeđu i gotovo sasvim crnu, ali to su samo najglavniji tonovi. Luschanova skala boje kože ima 36 brojeva, a još ćemo naći tonova, koji u toj skali nisu zastupani.
Ima bijele kože, koja uopće nema sposobnosti da stvara pigment, koja ni kod najjačeg sunčanja ne potamni, nego samo pocrveni, to je koža plavovlasih rasa, samo ta je koža uistinu bijela, odnosno ružičastobijela. Koža većine bijelih rasa ima pak sposobnost stvarati pod utjecajem aktivnih zraka pigment, pa može nakon duljeg sunčanja jako potamniti, ali većina tog pigmenta s vremenom opet iščezne, kada je sunčanje prestalo. Kod svih rasa osim evropske grupe, koža je stalno pigmentirana, kod tamnijih je rasa napose vezivni sloj natrpan pigmentnim stanicama, a ima ih i u potkožnom vezivu. Ne samo koža nego i sluznica na usnama i spolovilu pigmentirana je kod nekih Crnaca kao i sluznica u ustima. I bjeločnica sadržava tu pigment, tako da nije bijela, nego maslinastosmeđa.
Pigment u koži zaštita je protiv aktivnih zraka sunca, dok se nepigmentirana koža na suncu upali; dapače, može doći do ozbiljnih oštećenja. Koža Crnaca i drugih tamnih rasa prilagodba je, dakle, životu pod tropskim suncem, a da tijelo ne treba biti zaštićeno odijelom.
Kod novorođenčadi većine mongolidnih rasa nalazi se u kožnom vezivu na krstima pigmentna mrlja, koja kroz kožu modro prosijava. I kod bijelih rasa može se naći ta t. zv. mongolska mrlja, kako je Mayerhofer upozorio, vrlo često kod Dinarida. Ta mrlja kasnije obično iščezava.
Boja vlasi i dlaka ovisi kao i kod kože o vlastitoj boji rožne tvari kose i o pigmentu. U vlasima je uglavnom dvojak pigment, crvenkasti, koji je jednakomjerno proširen u kosi i proziran, i tamnosmeđi, koji se nalazi u malim zrncima. Količina jednog i drugog može biti vrlo različita. Kosa bez pigmenta bjelkasto je plava, od te boje vode dvije serije do sasvim tamne boje, jedna s vrlo malo crvenog pigmenta i druga s više crvenila. Prema količini tamnog pigmenta ima prva serija tonove od pepeljasto-plavog, pepeljastosmeđeg, mrkosmeđeg do mrkocrnog, a druga tonove od zlatno plavog, zlatno žutog, toplo smeđeg (kestenastog) do smeđe crnog. Ako nema tamnog pigmenta, nego samo crvenkasti, kosa je crvena. Crvena se kosa javlja kao anomalija kod rasa, koje imaju inače i tamni pigment u kosi, i to ne samo kod bijele rasne grupe, nego vrlo često i kod nekih crnih rasa u istočnoj Indiji. U odmaklo doba u srčiku vlasi ulazi zrak, koji bijelo prosijava, a može prestati i stvaranje pigmenta u vlasi, pa kosa posijedi odnosno pobijeli. I to je u velikoj mjeri nasljedno i rasno svojstvo, kao što i prerano ispadanje kose. Kod većine indijanskih rasa kosa uopće ne posijedi, dok kod mediterane rase biva vrlo rano sijeda i bijela, pa u području te rase često susrećemo ljude mladenačkog lica i bijele kose. Ćelavost značajna je za alpinsku rasu.
Kao što boja kose, ni boja šarenice nije ovisna o vanjskim utjecajima. Boja šarenice uzrokovana je bjelkastom bojom mreže vezivnih vlakana i vezivne sluzi u tkivu šarenice i pigmentnim stanicama, koje mogu biti raznoliko smještene u tom tkivu. Stražnji sloj šarenice krcat je pigmentom. Ako inače nema pigmenta u šarenici, prosijava crni sloj kroz prednji bjelkasti sloj, pa je šarenica modre boje, kao svaki mutan medij na crnoj pozadini (na pr. nebo ili dim). Ako ima pigmentnih stanica, kombinira se modra boja sa smeđim pigmentom, pa dolazi, prema položaju pigmentnih stanica, do različnih nijansa: zelenkaste, sive, sivosmeđe, svijetlosmeđe, tamnije smeđe do tamno, gotovo crno smeđe. Pigmentirane rase imaju pretežno tamno smeđu šarenicu.
Nedostatak svakog pigmenta, anomaliju, koja se javlja i kod različnih životinja kao mutacija (albinizam), nalazimo kao iznimnu pojavu kod svih ljudskih rasa. I među Crncima ima albina, a osobito su česti albini kod Indijanaca na Panamskom tjesnacu, ali nikada ne nastupa albinizam kao obilježje neke rasne skupine.
Nema sumnje, da postoje razlike između rasa i u pogledu nutarnjih organa, premda to dosele još nije dosta ispitivano. Razumljivo je, da se posebna pažnja posvećuje mozgu kao organu duševnih funkcija. Već je Tiedemann (1837) ispitao mozak Crnca i usporedio ga s mozgom bijelca i orangutana. No koliko su više ispitivani mozgovi različnih rasa, toliko se više činilo, da u tom pogledu nema rasnih razlika, jer se pazilo samo na opću građu i poredak moždanih zavoja. Prema novijim istraživanjima J. L. Shellsheara, koji je ispitao 44 mozga Australaca te ih usporedio sa 100 mozgova Kineza, ipak postoje rasne razlike. Mozak je Australca jednostavnije razbrazdan nego kineski, napose ima razlika u zatiljnom režnju, koji kod Australaca pokazuje primitivnije prilike. Za kakvoću mozga najvažnija je njegova citoarhitektonika, t. j. poređaj i gustoća stanica u različnim dijelovima moždane kore. U tom pravcu ima rezultata, u koliko se odnose na opseg vidnog polja kore (area striata na zatiljnom režnju), koje je kod Crnaca pronađeno većim nego kod Europida, uz to se kod ispitana 4 crnačka mozga 27% toga polja nalazi na vanjskoj površini mozga, a kod Europida tek 9,8%. Ovi početci pokazuju, da se može još mnogo očekivati od točnijeg ispitivanja citoarhtitektotektonike.
| Krvna tjelešca |
Krvni serum |
| |
0 |
A |
B |
AB |
| 0 |
— |
— |
— |
— |
| A |
+ |
— |
+ |
— |
| B |
+ |
+ |
– |
|
| AB |
+ |
+ |
+ |
— |
Ima važno fiziološko svojstvo čovjeka, koje je ovisno isključivo o nasljedstvu, te se ne može promijeniti nikakvim vanjskim utjecajem: to je kakvoća krvi s obzirom na neke osebine njenih bjelančevina, kako se javlja u krvnim grupama (v.). Radi se o sposobnosti seruma (sukrvice), da krvna tjelešca drugih ljudi aglutinira (sljepljuje u veće gomile) ili ne, i o svojstvu krvnih tjelešaca, da li ih serum drugih aglutinira ili ne. S obzirom na ta svojstva razlikujemo četiri vrste krvi, koje se označuju sa 0 (nula), A, B, i AB. Međusobni odnošaj tih vrsta krvi, ako miješamo serum jedne krvi s krvnim tjelešcima druge krvi, može se vidjeti iz gornje skrižaljke, u kojoj križić označuje aglutinaciju, a crtica, da nema aglutinacije.
Način nasljeđivanja tih svojstava poznat je u tančine. Kako je prvi pokazao Hirzsfeld, postotci su krvnih grupa kod različnih populacija veoma različiti, makar kod svake rase nalazimo sve četiri grupe. Kod pojedinca nam, dakle, njegova grupna pripadnost ne može kazati ništa, ali ako ispitamo nekoliko stotina ljudi neke populacije, već ćemo moći stvoriti (bez obzira na druga svojstva) neke zaključke o njihovoj rasnoj pripadnosti. Postotci krvnih grupa mogu napose poslužiti kao putokaz, ako se radi o pitanju srodstva nekih rasnih grupa. Tako je ispitivanjem krvnih grupa Cigana, koje su proveli Vérzar i Weszeczky, ustanovljeno, da se njihovi postotci poklapaju s postotcima krvnih grupa Indijaca, što potvrđuje indijsko podrijetlo Cigana. Jednako potvrđuju krvne grupe srodstvo Eskima i Indijanaca. Značajna je podjela krvne grupe B u Evropi, kojoj postotak od zapada prema istoku sve više raste. Svojstva, koja se odrazuju u krvnim grupama, ne odnose se samo na krv, nego se radi, kako je to napose Schiff pokazao, o svojstvima svih sličnih bjelančevina u tijelu. Neko naročito značenje tih svojstava za život organizma dosada nije uspjelo ustanoviti. Landsteiner, pronalazač svojstava krvnih grupa, i Lewine otkrili su još neka slična svojstva krvi, od kojih je ispitan udio faktora M i N kod različnih populacija. Radi se o svojstvu ljudskih krvnih tjelešaca, da budu aglutinirana od naročito priređenog seruma kunića. Krvna tjelešca mogu imati obadva svojstva, M i N, ili samo jedno od obadvaju. Postotci u nekim populacijama vide se iz tabele na str. 352.
Ima još daljnjih fizioloških rasnih razlika. Bilo ne kuca kod svih rasa jednakom brzinom; kod Evropljana prosječno 71—72 puta na minutu, kod Kirgiza 77, kod Tungida 65, kod Maja-Indijanaca samo 35—40 puta. Broj udisaja također nije jednak; dok Evropljanin udiše 16 puta u minuti, Tungidi 18—20 puta, Afganci 18 puta, a Kalmuci 21 put. Kožne žlijezde, znojnice i lojnice, raznoliko su razvijene kod pojedinih rasa. Osobito mnogo lojnih žlijezda imaju Australci, kojima je koža stalno masna i vrlo mekana, kao baršun. Crnci imaju vrlo mnogo znojnica, te se jakim znojenjem mogu bolje rashladiti nego bijelci. Melanezijci pak na Novoj Gvineji uopće se ne znoje. Izlučevine kožnih žlijezda, napose pazušnih, imaju svoj tipičan miris, koji je kod svake rase drukčiji, tako da govorimo o rasnom mirisu. Pripadnici iste rase ne osjećaju miris te rase, ali ga zato osjećaju pripadnici drugih rasa, obično vrlo neugodno. Taj miris djelomično ovisi o hrani, o vrsti masti, koja se upotrebljava, o različnim mirodijama i sl., ali i kod jednake hrane različite rase zadrže svoj specifičan miris. Osobito je neugodan za Evropljane miris andidnih Indijanaca i Crnaca, a Japanci se tuže, da im Evropljani zaudaraju. Isto su tako tvrdili francuski anamitski vojnici (Paleomongolidi), koji su poslije prvog svjetskog rata došli u Evropu, da im bijelci jako zaudaraju.
Izvjesne razlike postoje i u oštrini osjetila. Indijanci i Kinezi bolje osjećaju gorak okus nego bijelci, a Arapi manje. Oštrina vida također pokazuje razlike: ona iznosi kod Nijemaca 1,1, kod Rusa 1,4, Gruzina 1,6, Indijanaca 2,1, Nubijaca 3, a kod andidnih Indijanaca 5. Crnci mogu primijetiti do 10 puta manje kretnje u daljini, nego što ih bijelci jedva još primjećuju. Etiopidi imaju prema bijelcima smanjenu osjetljivost za crvenu i zelenu boju.
O nastupu spolne zrelosti kod različnih rasa ima mnogo podataka, napose o menarhi (početku menstruacije), za koju se prije mislilo, da se javlja to ranije, što je manja geografska širina. To oprovrgava već činjenica, da se kod Melanezijaka javlja tek oko 17. godine. B. Škerlj (1932) drži, da su, što se toga tiče, važni (barem u Evropi) ipak klimatski čimbenici, i to položaj siječanjske izoterme od 1°, kao i količina padalina i amplituda godišnje temperature. Ipak većina autora drži, da su i rasne razlike od značenja. To isto vrijedi i za starenje. Australidi počinju starjeti već poslije 40. godine, tako da oko 50. godine umiru od starosti. Kod žena nekih rasa, napose kod indidne, orijentalidne i polinezijske, opaža se, da rano sazriju, ali i rano venu, tako da počinju starjeti već s 20 godina, dok žene nordidne i nekih drugih rasa spolno sazrijevaju tek s 20 godina.
Ipak su fiziološka rasna svojstva slabo ispitana, pa znanost u tom pogledu ima još široko polje rada. Isto vrijedi za rasnu patologiju. Nesumnjivo se izvjesne bolesti kod nekih rasa češće javljaju nego kod drugih. Tako se navodi, da se šećerna bolest kod Židova javlja 6% puta češće nego kod autohtonih evropskih rasa. Kod Japanaca postoje neke nasljedne očne bolesti (Ogučijeva bolest), koje kod drugih rasa nisu poznate. Kod mnogih bolesti, koje se negdje češće javljaju nego drugdje, teško je odrediti, nijesu li tu po srijedi lih utjecaji klime i životnih uvjeta.
Kao što kod svih rasa nalazimo zastupane sve četiri krvne grupe, tako se slično kod svih rasa javljaju ona opća svojstva građe i reakcione sposobnosti tijela, koje zovemo konstitucijom (v.). Već su stari Indijci razlikovali tri tipa žene: gazelu, košutu i slonicu, što nije drugo nego Kretschmerovi leptosomni, atletski i piknički habitus. Francuzi (Sigaud, Chaillou, Mac Auliffe) razlikuju još četvrti, cerebralni (moždani) tip. U biti se radi o dva habitusa: leptosomnom, s uskim tijelom i dugim udovima, i eurisomnom, sa širokim bucmastim tijelom. Ostala dva habitusa samo su posebni oblici leptosomnoga odn. eurisomnoga. Od lubanja na str. 348 desno, prva pripada leptosomnom, a druga eurisomnom tipu. Ove konstitucione tipove nalazimo i kod životinja, napose kod domaćih, a naći ćemo ih na pr. i kod čimpanze. Radi se, dakle, o dvije vrste oblikovanja tijela, koje je čovjek imao već od početka svog postanka, a koje su se kod cijepanja na rase uvukle u sve pojedine rase, tek je njihova učestalost kod različitih rasa različita. Dinarska, nordijska, a jednako i nilotska rasa pokazuju mnogo češće leptosomni habitus, dok je eurisomni češći kod alpske, baltičke i paleomongolidne rase. I to su većim dijelom nasljedna obilježja, premda ovise mnogo i o izvanjim utjecajima, samo što nisu neposredna rasna obilježja, nego dva različna načina ispoljavanja rasnih osebina.
Budući da kao rasna obilježja dolaze u obzir samo nasljedna svojstva, vrlo je važno ustanoviti, u koliko se pojedina svojstva nasljeđuju i kojim se načinom prenose. Za to je pitanje od naročita značenja proučavanje križanaca između različnih rasa. E. Fischer (1913) ustanovio je na narodiću bastarda između Bura i Hotentota u Rehobotu u jz Africi, da se sva bitna rasna obilježja čovjeka prenose po Mendelovu zakonu, što znači, da su nasljedna. Slične su rezultate postigli Rodenwaldt (1928) na holandsko-indonezijskim križancima na otoku Kisar kod Timora i C. B. Davenport (1929) na križancima između njemačkih naseljenika i Crnaca na Jamaiki. Po ovim ispitivanjima kao i po istraživanjima rodoslovlja različnih porodica ustanovilo se, koja svojstva kod križanja prevladavaju (dominiraju), a koja su kod križanaca sakrivena (recesivna). To je od važnosti, ako želimo po somatskim osebinama miješane populacije prosuditi, koji je udio pojedinih rasa u toj populaciji.
Osim somatskih razlika, kako između pojedinih ljudi, tako i između rasa, svakako postoje razlike i u duševnim svojstvima, napose u temperamentu. Postoje li s obzirom na ona svojstva, koja se očituju u inteligenciji, među rasama kao cjelinama bitne razlike, o tome nema dosada takvih podataka, da bi mogli poslužiti kao podloga za sigurne znanstvene zaključke. Baš su u tome pogledu važni vanjski uplivi, što pokazuju iskustva na jednojajnim blizancima (dvojcima), koji su uzrasli u različitoj okolini i imali različiti odgoj.
Uz to treba kod prosuđivanja ljudskih rasa imati na umu, da je svaka rasa, kao i svaki organizam, što je bolje moguće, prilagođena životnim uvjetima svog područja. Razumljivo je stoga, da se ne će odmah snalaziti afrički Pigmejac ili cejlonski Veda, ako budu presađeni iz svoje prašume u evropski velegrad, a ne će bolje proći ni Evropejac, presađen u tropsku prašumu. Ako se svaka rasa promatra u vezi s okolinom, u kojoj živi, tada ne će biti govora o »nižim« ni o »višim« rasama, ni o manje ni o više vrijednima, jer je svaka rasa srasla sa svojom okolinom u harmoničku cjelinu.
IV. RAZVITAK NAUKE O LJUDSKIM RASAMA.
Već su sa staroegipatskih spomenika poznati likovni prikazi rasnih tipova, koji su tada živjeli u egipatskom carstvu, a pokazuju nam, da su Egipćani znali dobro opažati ljudska rasna obilježja.
Prvi pokusi, da se sustavno svrstaju ljudske rase, potječu od konca 17. st. Za te je pokuse svuda od najveće važnosti boja puti. Kod F. Berniera (1684), G. W. Leibniza, G. Buffona (1749) J. Huntera (1775) nastupaju pored tri glavne boje puti Laponci kao posebna rasna grupa. Svakako su ih tada bolje poznavali nego izvanevropske rase. U kasnijem izdanju svog djela (1789) Linné razlikuje 5 rasa: Homo albus (Evropejci, sjev. Afričani i Eskimi), H. badius (Mongoli), H. fuscus (Malajci), H. niger (Crnci) i H. cupreus (Indijanci). I. Kant (1775) pored bijelaca, hunske rase (Mongola), crnaca i Američana razlikuje još posebnu hindustansku rasu i »rasu sjeverne obale Azije« (Sibiridi). Boju puti obrazlaže Kant (u »Predavanjima o fizičkom zemljopisu«) utjecajem klime: pod utjecajem vlažne studeni nastali su plavi evropski sjevernjaci, suhe studeni bakreno crveni Indijanci, vlažne vrućine crnci i suhe vrućine maslinastožuti Indijci.
Od najtrajnijeg utjecaja bilo je djelo J. F. Blumenbaca De generis humani varietate nativa liber (1776). Kroz 150 godina vrijedila je njegova podjela rasa kao konačno ispravna, te je bila kao takva donedavna još sadržana u svim školskim knjigama. On razlikuje:
1. Kavkasku rasu, svijetle puti, crvenih obraza, crne ili smeđe kose, okrugle glave, ovalnog lica, uskog, lako ukrivljenog nosa i malih usta. Njoj pripadaju svi Evropljani osim Finaca i Laponaca, stanovnici sjev. Afrike, Azije do Oba, Kaspijskog mora i Gangesa. Na plavokose Evropejce ne uzima se naročiti obzir.
2. Mongolsku rasu, žuto smeđe puti, crne, tvrde kose, četverouglate glave, plosnatog lica, utisnutog nosa i uskog otvora vjeđa. Ostali stanovnici Azije osim južnih otoka, Laponci, Finci i Eskimi.
3. Etiopsku rasu, crne puti, crne kudrave kose, uske glave, kosozubu, s izbubrenim usnama, tupog nosa. Stanovnici Afrike osim sjevernog ruba.
4. Američku rasu, bakrene puti, crne tvrde kose, niskog čela, tupog nosa, širokog, ali ne plosnatog lica, s oštrim potezima. Urođenici Amerike osim Eskima.
5. Malajsku rasu. Smeđe puti, od svijetle mahagoni-boje do najtamnije kestenasto smeđe, s gustom crnom kudravom kosom, širokim nosom i velikim ustima. Stanovnici južnih otoka ili petog kontinenta sve do istočne Indije, s uključenjem pravih Malajaca.
O Australcima se ne izjašnjava pobliže, pa bi svakako pripadali ovoj petoj rasi. Da naziva bijelu rasu kavkaskom, koji je naziv, kakogod se činio zagonetan, ipak tada i kasnije općenito prihvaćen, ima svoj uzrok u pukom slučaju, da je Blumenbach imao u svojoj zbirci lubanju mlade Gruzinke, za koju kaže, da je tako lijepa lubanja, »da mora privući svako, za ovakvo što ikako osjetljivo oko«.
G. Cuvier (oko 1800), osnivač poredbene anatomije i oštar promatrač, bio je skromniji, istaknuo je samo ono, što je našao osnovano na tadašnjem znanju, pa razlikuje tri glavne rase: leukodermnu (bijelce), ksantodermnu (žutu) i melanodermnu (crnu). U tom krugu Blumenbacha i Cuviera kreće se većina kasnijih podjela prošlog stoljeća, samo su pojedine grupe djelomično pocijepali, bilo s geografskih, bilo s lingvističkih gledišta. Značajna su izlaganja J. Geoffroy-St. Hilairea (1863), koji ističe Hotentote i Bušmane kao posebnu glavnu grupu. Već su A. Desmoulins (1825), Bory de Saint Vincent (1863) i E. Haeckel (1868) odijelili Australce kao posebnu rasu, pored brojnih drugih rasa, koje razlikuju. Da su Australci jedna od temeljnih ljudskih rasa, jasno je istaknuo drugi veliki poredbeni anatom i oštar promatrač, T. H. Huxley (1870), koji razlikuje negroidnu, australoidnu, mongoloidnu, ksantokroidnu (plavokosi sjevernjaci) i melanokroidnu (tamnokosi bijelci) glavnu rasu, a uz to kao pododjele glavnih rasa 13 sekundarnih rasa. Tako se Huxley dosta približio prirodnoj genetskoj sistematici č-a.
Od velikog utjecaja bili su radovi švedskog antropologa A. Retziusa, koji je počeo 1842 svojim kraniometrijskim istraživanjima i dao poticaja za razlikovanje različnih tipova unutar evropskog pučanstva, ma da je to dovelo do toga, da su se precjenjivali rezultati mjerenja. Ta su mjerenja pokazala, da svako pučanstvo pokazuje najraznovrsnije mjere, pa da se uopće ne mogu izlučiti neke »čiste rase«, tako da je B. Topinard (1885) došao do zaključka, da ima samo različnih tipova, a da zapravo više nema pravih rasa, koje bismo mogli svrstati u genealoški sistem.
Zato je od najvećeg značenja djelo ruskog antropologa J. Denikera (1900), koji je radio u Parizu. On je na osnovu svih dotada sakupljenih morfoloških podataka pristupio sustavnoj podjeli ljudskih rasa. Ma da Deniker naglašava, da mu se čini, »da se ne može govoriti, kada se radi o ljudskom rodu, ni o vrsti, ni o varijetetu, ni o rasi u onom smislu, kao u zoologiji ili zootehnici«, to je ipak njegovo djelo postalo temeljem za noviju nauku o rasama, kakva se razvila upotrebom iskustava moderne nauke o nasljeđivanju, u nastojanju, da samo nasljedna svojstva budu uvažena kao rasna obilježja, pa da se tako dođe do prirodnog genealoškog sustava ljudskih rasa. Kao najvažnije osebine za sustavnu podjelu upotrebljava Deniker: boju puti, boju šarenice, boju i oblik kose, oblik nosa, oblik glave i visinu tijela. On razlikuje 29 rasa, koje grupira u 17 skupina, i to:
I. 1. Bušmanska rasa (Bušmani i Hotentoti).
II. Negroidna grupa: 2. Negriti (afrički i indijski Pigmejci).
3. Crnci. 4. Melanezijci (Nova Gvineja i susjedni otoci).
III. 5. Etiopska rasa (Somalija, Abesinija).
IV. 6. Australska rasa.
V. 7. Dravidska rasa (ist. Indija).
VI. 8. Asiroidna rasa (dio Armenaca, Perzijanaca, Kurda i Židova). VII. 9. Indo-afganska rasa.
VIII. Sjevero-afrička grupa. 10. Arapska ili semitska rasa. 11. Berberi.
IX. Melanokroidna grupa (Tamnokosi Evropejci): 12. Litoralna ili Atlanto-mediteranska rasa. 13. Ibero-inzularna rasa (ove dvije danas se označuju kao mediteranska rasa). 14. Zapadna rasa (današnja alpska). 15. Jadranska ili dinarska rasa.
X. Ksantokroidna grupa (plavokosi Evropejci); 16. Nordijska i 17. orijentalna rasa (istočno-baltijska).
XI. 18. Ainu (na Sahalinu i u sjev. Japanu).
XII. Oceanska grupa: 19. Polinezijska rasa (Havai, Samoa, Nova Zelandija). 20. Indonezijska rasa (Sundski otoci).
XIII. Američka grupa: 21. Sjevero-američka, 22. Srednjo-američka, 23. Južno-američka i 24. Patagonska rasa.
XIV. 25. Eskimska rasa.
XV. 26. Laponska rasa.
XVI. Evrazijska grupa: 27% Ugrijska rasa (istočni Finci, Samojedi Jakuti). 28. Turkotatarska rasa.
XVII. 29. Mongolska rasa (Mandžurija, Korea, Kina).
Grupacija je provedena isključivo na osnovi somatskih svojstava bez obzira na etničku i lingvističku podjelu, makar se za rase još često upotrebljava etničko nazivlje. Deniker je bio prvi, koji je istaknuo naročitu dinarsku rasu.
Poslije Denikera pokušali su sustavno razvrstati ljudske rase, a da kod toga upotrijebe latinsko nazivlje u smislu zoološke nomenklature, L. Wilser (1912). V. Giuffrida-Ruggeri i G. Sergi (1911). Wilser i Sergi uključuju u svoje sustave i sve fosilne forme; obadva razlikuju više prirodnih vrsta. Pored fosilnih rodova i vrsta Wilser govori o tri sadašnje vrste čovjeka, Homo niger (Crnci), H. brachycephalus (svi Mongoli i brahicefalni bijelci) i H. europaeus (mediteranska i nordijska rasa), svaka s brojnim varijetetima. Po Wilseru bila bi, dakle, većina evropskog pučanstva križanci dviju vrsta čovjeka Sergijev sustav razlikuje 5 rodova (→ Podrijetlo čovjeka str. 242), 2 izumrla i 3 živa više vrsta: Notanthropus (grč. νότος jug«) eurafricus (dolihocefalni bijelci, Etiopljani, Polinezijci, Dravide, Ainu), N. afer (Crnci), N. australis (Hotentoti); Heoanthropus (grč. ἔως »istok«) eurasicus (brahicefalni bijelci), H. arcticus (Laponci, Sibiridi), H. orientalis (Mongoli); Hesperanthropus (grč. ἑσπέρα »zapad«) Columbi (Indijanci, Eskimi), H. patagonicus. Svaka je vrsta s više varijeteta, a neke su od ovih podijeljene i na subvarijetete. Kako se vidi, Sergi pribraja različnim rodovima i one rase, koje sačinjavaju evropsko pučanstvo. Sergijev sustav polazi s gledišta, da su dolihocefalni Evropejci u bližoj genealoškoj vezi s Crncima i Hotentotima, tako da bi bila svojstva, koja su im zajednička s brahicefalnim bijelim rasama, samo posljedica konvergentnog razvitka, pa je razumljivo, da taj sustav nije našao mnogo pristaša.
| Subspecies (podvrsta) |
Serija |
Varietas (rasa) |
Subvarietas (podrasa) |
| Homo sapiens albus Imelin Europidi |
Plava rasna zona |
Nordidi |
Teutonordidi Dalofalci, Fenonordidi |
| 1st. Europidi |
- |
| Smeđa rasna zona |
Mediteranid! |
Gracilni, Eurafrikanidi. Saharidi |
| Orijentalidi |
- |
| Indidi |
Gracilni, Sjeverni, Indobrahidi |
| Polinezidi |
Polinezidi, Mikronezidi |
| Brdska rasna zona |
Alpinci |
Zapadni Alpinci, Lapidi |
| Dinaridi |
- |
| Armenidi |
- |
| Turanidi |
Aralidi, Pamiridi |
| Staroeuropidi |
Vedidi |
Veda, Gondidi, Malidi, Toalidi, Senoidi |
| Ainuidi |
- |
| Homo sapiens leiotrichus Bory |
Mongolidi |
Polarna rasna zona |
Sibiridi |
Zapadni, istočni |
| Eskimidi |
- |
| Istočni Mongolidi |
Tungidi |
- |
| Sinidi |
Sjeverni, srednji, južni |
| Paleomongolidi |
Palaungidi, Šanidi, Protomalajidi, Deuteromalajidi |
| Koisanidi |
Koisanidi |
Koidi, Sanidi |
| Amerikanidi |
Sjeverni Indijanidi |
Pacifidi |
- |
| Centralidi |
- |
| Silvidi |
- |
| Margidi |
- |
| Južni Indijanidi |
Andidi |
- |
| Patagonidi |
- |
| Brazilidi |
- |
| Lagidi |
- |
| Fuegidi |
- |
| Homo sapiens afer L Negridi |
Kontaktna rasna zona |
Etiopidi |
Sjeverni, središnji, južni |
| Indomelanidi |
Sjeverni, južni |
| Zapadni Negridi |
Sudanidi |
- |
| Nilotidi |
- |
| Kafridi |
- |
| Paleonegridi |
- |
| Istočni Negridi |
Neomelanezidi |
- |
| Paleomelanez. |
- |
| Australidi |
- |
| Pigmidi |
Bambutidi |
- |
| Negritidi |
Aetidi, Semangidi, Andamanidi |
Sistem ljudskih rasa po Eickstedtu (1937)
Giuffrida-Ruggeri nabraja 8 glavnih rasa: 1. Homo sapiens australis, 2. H. s. pygmaeus, 3. H. s. indoafricanus (Dravida i Etiopljani). 4. H. s. niger, 5. H. s. americanus, 6. H. s. asiaticus, 7. H. s. oceanicus (Polinezijci, Ainu), 8. H. s. indoeuropaeus (evropski, sjeveroafrički i južnoazijski bijelci). Ovomu se sustavu s morfološkog gledišta ne može mnogo prigovoriti.
Montandonov sustav (1928) predočuje drugu mogućnost primjene morfoloških značajaka. Montandon razlikuje 20 rasa, koje su poredane u 5 grupa:
A. Pigmoidna skupina sa steatopignom (Hotentoti i Bušmani) i pigmejskom rasom.
B. Negroidna skupina s tasmanskom, papuanskom, negridskom, etiopskom i dravidskom rasom.
C. Vedoaustralska skupina s Veda i australskom rasom.
D. Mongoloidna skupina s paleoamerindnom, neoamerindnom (obadvije Indijanci), eskimskom, paleosibirskom, mongolskom i turanskom (Tatari, Turci) rasom.
E. Evropidna skupina s laponskom, Ainu, plavom, alpsko-armenskom i smeđom rasom (potonja bi obuhvatala dolihocefalne tamnokose bijelce).
Najznatniji pokušaj, da se ljudske rase poredaju u genealoški sustav, potječe od E. Fischera (1932). Velike poteškoće za sistematiku rasa činile su osebine, koje nalazimo slično razvijene kod inače različitih rasa, na pr. orlovski nos kod Dinarida, kod Kirgiza, kod amer. Indijanaca, ili spiralna kosa kod Crnaca i kod Tasmanaca, visoki rast kod brojnih, inače vrlo različnih rasa. Zbog toga je bilo različnih mišljenja, koje zajedničke osebine treba shvaćati kao izraz zajedničkoga podrijetla, a koje kao konvergencije sporednog značenja. S obzirom na eksperimentalna genetička iskustva drži Fischer, da nasljedni čimbenici čovjeka naginju k tome, da lakše mutiraju u izvjesnim pravcima nego u drugima. Prisutnost pojedinačnih sličnih ili jednakih svojstava kod inače različitih rasa tumači stoga Fischer tako, da su se javile neovisno iste mutacije kod različnih loza, pa da takva obilježja ne znače neku genealošku vezu nego samo konvergenciju. To je svoje shvaćanje Fischer prikazao u rodoslovnim crtežima ljudskih rasa, u kojima je naznačio, kada su se javile pojedine mutacije, koje su dovele do odvajanja različnih loza.
Kao podlogu svom rodoslovlju uzima Fischer gledište, koje je već i prije istaknuo, da postoje 4 glavne grupe ljudskoga roda, australska, crnačka, mongolska i evropidna, i da Australci imaju u morfološkom pogledu svoj položaj na korijenu sadašnjeg ljudskog roda. Razumije se, da ima ovaj prvi, na temelju pogleda moderne genetike provedeni genealoški sustav, još mnogo što hipotetsko, ali će svakako biti teško prigovoriti bitnim njegovim smjernicama.
U pravcu Fischerovih nazora kreće se i Weinert kod svog pokusa, da prikaže postanak ljudskih rasa. On razlikuje tri razvojne loze, srednju liniju bez naročito specijaliziranih obilježja i dvije postrane, svaku specijaliziranu u drugom pravcu. Na bazi srednje linije, koju nadovezuje na Neandertalca, stavlja Australce, a na gornjem kraju završava srednja linija s Evropidima. Indonezijce, kojih Fischer ne navodi u svom rodoslovlju, drži Weinert za proizvod križanja australidnih i mongolidnih elemenata. Što se tiče Laponaca, slaže se s Eickstedtom, da su pripadnici Evropida i da potječu od onih prvobitnih Alpida, koji su već u davnini odgurnuti na sjever.
Najmoderniji prikaz ljudskih rasa potječe od E. Eickstedta, izvrsnog poznavaoca stanovništva istočne Indije, koji je unio mnogo izvornih misli u promatranje ljudskih rasa u vezi s geografskim čimbenicima njihova razvitka. Eickstedt je sakupio i kritički obradio svu građu, koju je pribrala antropološka znanost o ljudskim rasama, da svakoj što jasnije odredi njezino mjesto i njezinu značajku, tako da njegovo djelo pruža zaokruženu sliku sadašnjeg stanja nauke o rasama. Eickstedt je i dosljedno proveo nazivlje rasa. Dugi latinski nazivi od tri do četiri riječi, kako ih je predložio Sergi, nezgrapni su. Nije ispravno upotrebljava i etničke ili, dapače, lingvističke nazive za rase, jer se time stvara zabuna, pa se rase nekako poistovjetuju s pripadni cima nekog naroda ili neke jezične grupe. Ta rasa je isključivo biološki pojam. Zato Eickstedt upotrebljava dosljedno za rase bilo geografske, bilo već inače udomljene oznake, ali sa završetkom -id, koji je način već prije katkada upotrebljavao napose Fischer, na pr. Dinaridi, Eskimidi, Orijentalidi. To su kratke i precizne oznake, lako upotrebljive u govoru i pismu, koje ne dopuštaju nikakve sumnje, da se radi samo o rasi, a ne o geografskom, etničkom ili lingvističkom pojmu. Ovo se nazivlje sve više prihvaća, jer je jasno i praktično.
Eickstedt govori o tri glavne podvrste (subspecies): europidnoj, mongolidno-amerikanidnoj i negridnoj. Svaku od njih dijeli na više serija, a te na rase, neke rase pak još na podrase, kako to predočuje tabela na str. 353.
Rasna teorija.
Neovisno o znanstvenoj nauci o rasama razvilo se koncem prošlog stoljeća naučavanje, koje nazivaju rasnom teorijom ili rasizmom. Bit je rasizma uvjerenje, da je neka rasa najbolja i najsposobnija, da po duševnim sposobnostima natkriljuje ostale rase, pa da je stoga »od Boga izabrana« da upravlja svijetom i sudbinom ostalih rasa. Ima različnih rasizama, a napose postoji velika literatura o nordijskom rasizmu, koji je osnovao francuski povjesničar J. A. Gobineau (1853), a dalje su ga izradili J. Vacher de Lapouge (1899), H. S. Chamberlain (1899) i dr. Promatrajući povijesni razvitak kultura indoevropskih naroda Gobineau nalazi, da su stvaranju tih kultura vidan dio pridonijeli ljudi one rase, koju danas zovemo nordijskom. Ova je konstatacija, doduše, vrlo vjerojatna, ali zaključak, da je ta rasa jedina sposobna za stvaralački polet, ovom konstatacijom nikako još nije logički osnovan. Tom konstatacijom, naime, nije dokazano, da i druge rase ne bi mogle postići pod prikladnim uvjetima slične uspjehe i razmaknuti se u stvaralačkom poletu. Sigurno je, da ima među rasama razlika kako u somatskim, tako i u duševnim obilježjima, makar o potonjem pitanju nema mnogo eksaktnih podataka, jer je baš kod duševnih svojstava teško izlučiti utjecaj vanjskih čimbenika. Ali ako i ima razlika između rasa u njihovu duševnom stilu, time još nije kazano, da je neka rasa vrednija od druge. Svakome će biti duševni stil njegove rase subjektivno najbliži, najvredniji, pa će ga ljudi njegove rase privlačiti, tako da će se između ljudi jednake rase lakše razviti društvovne veze nego između ljudi tuđih po rasi. Ali takvi subjektivni momenti ne mogu biti podlogom za određivanje objektivne vrednote rasa. Pa eksaktna znanost uopće i ne pozna određivanje vrednote, nego konstatira postojeće pojave i istražuje njihove uzroke.
Rasizam teži za tim, da se u korist napretka kulture sačuva što čišća ona rasa, koju smatra najsposobnijom. Ima, međutim, činjenica, koje bi se mogle i u suprotnom smislu tumačiti; da naime baš miješanje nekih rasa pospješuje razvitak onih duševnih snaga, koje dovode kulturnom napretku. I E. Fischer se jednom izjavio u tom smislu, a napose zastupa to gledište Kretschmer. Gerlach je sakupljanjem podataka o mjestima rođenja istaknutih kulturnih radnika došao do zaključka, da se u Njemačkoj u svako doba rađalo najviše pjesnika i slikara u onoj zoni, gdje su se miješali Nordidi s Alpidima i Dinaridima. Ovo još nije neki dokaz, da je stvaralačka snaga onih kulturnih radnika bila posljedica baš miješanja rasa, jer ima i drugih stvari, koje bi mogle doći u obzir, ali svakako one činjenice dopuštaju Gerlachovo tumačenje, i nikako ne govore u prilog »čiste rase«. Uistinu, može se često ustanoviti, da su istaknuti ljudi miješane rase. O. Ammon (1893) dolazi, doduše, na osnovu antropološkog ispitivanja 30 članova nekog znanstvenog društva u Karlsruhe do zaključka, da su »u velikoj mjeri dolihocefalni svi učenjaci, koji su postigli znatnih uspjeha«. Ako tu tvrdnju, na koju Lapouge nadovezuje dalekosežne zaključke, ispitamo na stvarnim činjenicama, lako ćemo se uvjeriti, da se radi kod Ammona o prenagljenoj generalizaciji slučajnog nalaza. Ta ima podataka o nekim antropološkim obilježjima najvećih umova i stvaralačkih duhova, iz kojih proizlazi, da mnogi od njih nisu imali nordidna obilježja, makar su potjecali iz djelomično nordidnih područja. Lubanja veleuma G. W. Leibniza, koju je istražio W. Krause, jest ultrabrahicefalna, s indeksom 90,3, što odgovara indeksu 92 u životu. Drugi veleum, I. Kant, imao je indeks glave od 90, bio je dakle hiperbrahicefalan, kako su to utvrdili Kupffer i Bessel-Hagen; indeks njegove lubanje iznosi 88,5. Sadreni odljevak glave A. Schopenhauera, koji je opisao Gwinner, pokazuje mjere, po kojima se može, kako ističe Weidenreich, izračunati indeks od 90, bio je dakle također hiperbrahicefalan. Za Beethovena je poznato, da je imao crnu kosu, a lice mu je bilo tipično alpidno. Goetheov šešir pokazuje, da mu se kretao indeks glave oko 88, bio je hiperbrahicefalan; kosa mu je bila smeđa, isto tako i šarenica, kako je utvrđeno po slikama. Put mu je bila žućkasta, te je na suncu smeđe potamnila, a lice mu odaje mnogo dinaridnih crta.
Iz ovakvih se i sličnih primjera vidi, da smo daleko od toga, da bismo mogli sa somatskim rasnim svojstvima neposredno vezati neke specifične duševne osebine i sposobnosti, pa da rasistička nastojanja nemaju oslona u znanosti o rasama.
V. PODJELA I OPIS LJUDSKIH RASA.
Sustavna podjela ljudskih rasa ili antropografija mora nastojati, da kao biološka nauka prikazuje rase po načelima, koja su inače mjerodavna kod sustavnog prikazivanja živih organizama, naime u prirodnom sustavu, u kojem su manje skupine prema svom srodstvu udružene u veće, a ove svrstane u morfološkom redosljedu, u kojem se odrazuju pravci prirodnog razvitka. Gotovo svi antropolozi slažu se u tome, da postoje 3 glavne rasne grane, bijela, žuta i crna, a da ih veže na korijenu australska grupa, koja je najbliža pračovjeku. Ali ima i jedna grupa, kojoj je teško odrediti položaj u jednoj od tih grana, s različnim vlastitim obilježjima; to su Koisanidi, koje je već J. Geoffroy-St. Hilaire istaknuo kao posebnu skupinu. Većina je, doduše, pribraja Crncima, Eickstedt pak Mongolima. Pa i Eskimidi i Indijanidi čine jasno ograničene skupine. Svi se slažu u tome, da su se ove dvije grupe razvile iz Mongolida, pa kako ni u zoološkom sustavu poznavanje podrijetla neke skupine nije zaprekom, da bude istaknuta kao posebna skupina, ako je oštro ograničena, tako možemo dosljedno i Eskimide i Indijanide smatrati posebnim skupinama, ma da tim nikako ne poričemo njihov razvitak iz prvobitnih Mongolida.
Na temelju izloženoga mogli bismo u svrhu sustavnog prikaza razlikovati slijedeće rasne skupine: 1. australidnu, 2. negridnu, 3. koisanidnu, 4. mongolidnu, 5. eskimidnu, 6. indijanidnu i 7. evropidnu, a inače ćemo se držati Eickstedtovih sistematskih jedinica.
 tabla I.
tabla I.Ova skupina obuhvaća rase, koje stoje po svojim somatskim svojstvima najbliže pračovjeku, napose to vrijedi za urođenike Australije, Tasmanije, neke urođenike Nove Gvineje i za Vedide. Put im je tamna, većinom gotovo crna i baš kod najprimitivnijih kosa je gusta i valovita. Ima i patuljastih forma, a kod nekih je kosa spiralno kovrčasta.
Australidi (tab. I. sl. 1 i 2)
Prosječne mjere: visina 167—171,5 cm, indeks glave 71,7—74,7. Stas je vitak, noge relativno duge, čelo plitko, iznad očiju nalaze se obično jake nadočne nabubrine, nos je povišen, ali širok, čeljusti kosozube, kosti lubanje debele i masivne. Sjedalo je slabo razvijeno Palac je na nozi jako pokretan. Put je tamno smeđe crna, kosa tamno smeđa, valovita ili kovrčasta. Kod djece je katkada kosa svijetla poput lana. Kod muškaraca su jako razvijeni brada i brkovi. Unatoč toliko primitivnih obilježja Australidi ipak pokazuju neke evropidne crte: prominentan nos, zaobljeno lice, oblik kose, bradu i brkove. S druge strane od svih živih rasa najsličniji su pračovjeku, tako reći živi fosili. Prvobitno prošireni čitavim australskim kopnom danas su potisnuti u neplodne središnje i sjeverne krajeve.
Paleomelanezidi (tab. I. sl. 3).
Prosječne mjere: visina 166,4—167,7, indeks glave 72,5—76,5. Tijelo je zdepasto, ruke dugačke, čelo obično ravno i natkriljuje oči jakim nadočnim lukovima, oko usana lice je mesnato, tako da usne strše naprijed. Nos je nizak, konkavan i vrlo širok, indeks nosa od 92—100. Put jako tamna, ali s nešto crvenkaste primjese. Usne tamno ljubičaste, kosa crnosmeđa, spiralno kudrava. Katkada se javlja crvena kosa. Dlakavost prilično razvijena, isto tako brada i brkovi. Najgrublje crte nalazimo u Novoj Kaledoniji, nešto ublaženi tipovi živu na Novim Hebridima i Salamonskim otocima. Tragova Paleomelanezida ima na Novoj Gvineji i otocima južno odatle. Oni su tvorili i prvobitno stanovništvo Nove Zelandije, ali su ih u historijsko vrijeme evropidni Maori većim dijelom istrijebili te se s ostatcima pomiješali.
Tasmanidi (tab. I. sl. 4).
Prosječne mjere: visina 166 cm, indeks glave 76—77. Ove bivše stanovnike otoka Tasmanije smatraju neki kao H. Basedow, R. Biasutti i E. Fischer posebnom rasom, a drugi, među njima Deniker i Eickstedt, samo južnim odvojkom Paleomelanezida. Na žalost istrijebili su ih engleski kolonisti u toku od 70 godina. Zadnja Tasmanka, Trukanini, umrla je g. 1876. Lice je Tasmanida bilo plosnato s debelim usnama i najširim nosom, što postoji kod čovjeka. Uho veliko. Put je bila tamno smeđa, kosa gusta, spiralno kudrava, dlakavost tijela jaka s gustom bradom kod muškarca.
Neomelanezidi (tab. I. sl. 5).
Prosječne mjere: visina 158—161,8, indeks glave 73,5—80,3. Tijelo je čvrsto i vitko, noge dugačke. Lice je ovalno, obrazi zaobljeni kao kod Evropida. Čelo je usko i visoko. Nadočni lukovi nisu naglašeni, usne su pune. Nos je, doduše, širok (indeks 85—88), obično ravan i prominentan, često ukrivljen, kao t. zv. židovski nos. Put je jako tamna, smeđe crna, kosa je obično tamno smeđa, gusta i spiralno kudrava kao vlasulja. Dlakavost je neznatna, brada rijetka. Često je kosa crvena, dapače, crveno plava, ili miješana, sa smeđim i crveno plavim pramenima. Spolna se zrelost javlja kod Neomelanezida kasnije nego kod Evropida, jasan znak, da se radi o rasnom svojstvu, a ne o nekim klimatskim utjecajima. Neomelanezidi sačinjavaju glavni dio stanovnika Nove Gvineje, ali im ima tragova i na okolnim manjim otocima.
Azijski Pigmidi ili Negritidi (tab. I. sl. 6).
Prosječne mjere: visina 146,8—148,1 cm, indeks glave 81,4—83,5. Makar se radi o patuljcima, proporcije su njihova tijela jednake kao kod visokih rasa, za razliku od afričkih Pigmeja, kojih tijelo pokazuje dječje proporcije. Glava je okrugla, lice plosnato, nos širok i nizak. Usne su debele, ali nisu izbubrene. Kosa je spiralno kudrava, često fil-fil-tipa. Lakša je steatopigija česta. Negritide nalazimo na Andamanskim otocima (Andamanidi), gdje su najčišći, na Malaki, (Semangidi), ali već miješane s mongolidnim elementima i na Filipinima (Aetidi), gdje su također već miješani s ostalim pučanstvom.
Vedidi (tab. I. sl. 7—9).
Prosječne mjere: visina 154—159,7 cm, indeks glave 73,3—78. Makar su Vedidi vrlo primitivni, ipak pokazuju toliko evropidnih obilježja, da bismo ih mogli nazvati nekim prijelazom između Australida i Evropida. Eickstedt ih i pribraja Evropidima. Vedidi imaju obilježja, koja se kod nas ističu kod djece. Okruglo lice, visoko čelo, mali frnjasti nos, donekle dječje proporcije tijela s nježnom šakom i stopalom. Eickstedt (1937) razlikuje pet tipova. Najprimitivniji su Malidi u Indiji, s debelim usnama, širokim licem, smeđecrnom puti i gustom kovrčastom kosom. Njima su slični pripadnici plemena Veda na Cejlonu, koji su pak već mnogo miješani sa Singalezima. Gondidi, koji živu na Dekanskoj visoravni, viši su po stasu, imadu uži i ravan nos te mnogo svjetliju put. Toalidi su ostatci Vedida na sundskim otocima, gdje su već prilično pomiješani s Paleomongolidima; isto vrijedi za peti tip, Senoide na Malaki, u Siamu i Burmi. Vedidi su bez sumnje prvobitni prastanovnici prednje i stražnje Indije, koji su od kasnijih useljenika potisnuti u šumovite predjele ili u džungle kao na Cejlonu, gdje provode skroman život prezreni od Indijaca, koji vide u njima »neku vrstu indijskih životinja«. Uistinu, to su skromni, čestiti i sretni ljudi, o kojima govore svi istraživači simpatično.
Indomelanidi (tab. I. sl. 10).
Prosječne mjere: visina 159,4—162,9, indeks glave 76,2. Ovim imenom označuje Eickstedt crnu rasu u Indiji, koja bi uglavnom odgovarala dravidskoj ili melanoindijskoj rasi Denikerovoj. Lice je pravokutno, čelo uspravno, nos ravan, ali širok, usne pune. Put tamno smeđa, kosa lako kovrčasta. Svakako, Indomelanidi nisu neka prvobitna rasa, nego proizvod križanja tamnoputih elemenata s evropidnom indskom rasom, pa bi bili nešto analogno s Etiopidima u istočnoj Africi. Pretežno su melanidne rase Tamili u pokrajini Madras i pleme Munda na sjeveroistoku Dekana, ali tu ima mnogo gondidne primjese.
2. Negridna skupina.
Kako već ime ove skupine kazuje, glavno joj je obilježje tamna put, koja je kod najtipičnijih predstavnika crna sa sivkastim tonom. Kosa naginje spiralnom kovrčanju. Dlakavost je slaba, a tako i terminalne dlake, brada i brkovi. Koža je nježna i jako glatka. Lice je slabo profilirano, s niskim, širokim nosom, bez izrazitog bradnog šiljka. Kratko tijelo često je zdepasto, s dugim udovima, napose su noge duge i vitke, jer su mišići golijeni slabo razvijeni. Šaka je gracilna, a stopalo veliko. Negridi su isključivo stanovnici Afrike, gdje se valjda stvorila ta rasna grupa. Dugo se za Crnce držalo, da su jedinstvena rasa. I E. Fischer drži, da su razlike nastale tek u kasnije vrijeme. Montandon (1928) se prvi izjasnio, da treba razlikovati središnji blok Crnaca afričkih šuma i skupinu afričkih stepa, koje su dalje diferencirane i sadržavaju manje ili više evropidne primjese. Podjela Negrida pokazuje prema tome četiri pojasa, kojima je poredak posljedica pritiska evropidnih elemenata sa sjeveroistoka.
Paleonegridi (tab. I. sl. 11).
Prosječne mjere: visina 157—166 cm, indeks glave 74—78. Na Kongu je stas niži i glava okruglija, a u Gvineji je tijelo više i glava dulja. Nos plosnat, sedlast, s vrškom, koji iskače kao gumb. Čeljusti su kosozube, usne debele, nabubrene. Lične se kosti jako ističu. Lice je nisko i široko sa zaokruženim čelom. Uho je maleno i stoji koso. Put je tamno smeđa, na suncu biva gotovo crna. Usne su ljubičaste boje, sluznica u ustima je pigmentirana. Polumjesec na noktima je modrikasto crven, svojstvo, koje se tvrdokorno nasljeđuje i kod križanaca. Kosa je crna, kratka i gusto kudrava, često fil-fil-oblika. Spolna zrelost nastupa kod žena već s 15 godina, a sa 25 godina već su uvele. Paleonegridi živu u afričkim prašumama Konga, u francuskoj ekvatorijalnoj Africi, Angoli i Gvinejskim obalama sve do Senegambije, gdje dolazi do miješanja sa Sudanidima.
Bambutidi ili afrički Pigmidi (tab. I. sl. 12).
Prosječne mjere; visina 141—143 cm, indeks glave 75,6—79,4. Ti afrički patuljci pokazuju infantilne proporcije tijela, s razmjerno velikom glavom i kratkim nogama. Čelo je izbočeno, ali nisko, lice zaobljeno. Nos je vrlo širok, ali viši nego inače kod Crnaca. Usne nisu nabubrene, dapače mogu biti tanke. Koža je smeđa s crvenkastim tonom, dakle razmjerno svijetla. Kosa je smeđe crna i kudrava u obliku fil-fil. Bambutidi sa svojim različnim plemenima Aka, Bata, Efe, Babongo i dr. živu u malim zajednicama u najdubljim prašumama Konga i sežu sve do Abesinije. Makar su prezreni od drugi Crnaca, ipak dolazi do križanja s drugim crnačkim rasama.
Kafridi, nazvani i Bantuidi (tab. I. sl. 13 i 14).
Prosječne mjere: visina 165,7—169,6 cm, indeks glave 74,0–77,0. Tijelo je čvrsto, kod muškaraca često atletske konstitucije. Donji dio kralježnice ukrivljen je natrag, tako da krstača stoji u kosom položaju. Kod žena je trup obično dugačak i noge kratke. Lice je kod muškaraca uglato, kod žena okruglo, čelo izbočeno. Čeljust pokazuje lako izražen bradni šiljak. Nos je plosnat s napuhnutim krilima. Usne su debele, nešto nabubrene, poduprte od umjerene prognatije. Koža ima različite tonove, od žuto smeđeg do tamno smeđeg. Šarenica je smeđa, kosa je spiralno kudrava, često tipa fil-fil. Dlakavost je tijela slabo razvijena, isto tako obrve; muškarci imaju rijetku bradu. Boja je kose gotovo crna, samo kod katkada crvenkasto smeđa Kod nekih skupina ima i evropskih napose kod Herera i Zulukafra. Kafridi živu u istočnoj i južnoj Africi. Njihovo su obitavalište stepe i savane, ali su prodrli i duboko u prašume.
Stanovnici Madagaskara bili su prvobitno ove rase, ali su poslije dobili mnogo mongolske primjese, koja tamo nije jednakomjerno proširena. Dok kod Sakalava prevladava kafridni elemenat, Hove su pretežno mongolidni.
Sudanidi (tab. I. sl. 15).
Prosječne mjere: visina 168,2—175,2 cm, indeks glave 72,6—79,4. Kod Sudanida su tipično crnačke osebine najjače razvijene. Tijelo je još čvršće nego kod Kafrida, širokih ramena, s opsežnim grudnim košem, evropidnih proporcija, samo što su podlaktica i golijen nešto dulje. Mišići golijeni bolje su razvijeni nego kod drugih Negrida. Gležnjevi su uski, ruke uske i nježne. Čelo je usko, tako da je lice preko obraza najšire. Nos je konkavan s vrlo širokim napuhnutim krilima. Usne su jako debele i izbubrene, te strše uslijed kosozubih čeljusti naprijed. Put je tamno smeđo crna, usne su ljubičaste ili modro sive, kratka crno smeđa kosa gusto je kudrava, ali rijetko postizava fil-fil-tip.
Centar je Sudanida zapadni Sudan, Senegambija i Nigerija. Ašanti, dahomejski i beninski Crnci pretežno su sudanidne rase.
Nilotidi (tab. I. sl. 16).
Prosječne mjere: visina 172,8—182,2 cm, indeks glave 71,7—79,7. Nilotidi idu među rase najvišeg stasa, visina tijela preko 2 m nije nikakva rijetkost. Tijelo je vitko i čvrsto, noge su razmjerno jako dugačke, mišići golijeni slabo su razvijeni. Lice ima mnogo evropidnih crta. Nos je na korijenu lako povišen s ravnim hrptom, ali su krila napuhnuta. Usta su malena, a usne nisu jako izbubrene. Čeljusti su ravnozube, donja čeljust ima dobro razvijen bradni šiljak. Lice je ovalno. Put je gotovo crna, najtamnija od svih rasa. Crna je kosa gusto kudrava, često fil-fil-tipa. Dlakavost je slabo razvijena. Područje ove rase nalazi se oko gornjeg Bijelog Nila. Plemena Dinka, Nuer, Atvot, Šiluk, Džaluo, Ačoli, Lango, Sara i dr. pretežno su nilotidne rase.
 tabla II.
tabla II.Etiopidi (tab. II. sl. 1 i 2).
Prosječne mjere: visina u sjevernom području 163,6—167,4, u južnom 170—180 cm; indeks glave 72,0—76,1. Tip Etiopa jasno odaje, da se ne radi o prvobitno jedinstvenoj rasi, nego o križancima Negrida i Evropida, i to Orijentalida. Stoga im E. Fischer i drugi ne priznavaju karakter zasebne rase. Ali su obilježja Etiopida već tako izjednačena, a razlikuju ih od drugih skupina, da potpuno odgovadaju definiciji rase. Prije je ova rasa često nazivana hamitskom s obzirom na jezike, koji su ondje običajni. Etiopidi su čvrstog, visokog, vitkog tijela, evropidnih proporcija. Lice je dugoljasto, lijepo profilirano, s uskim visokim nosom, samo što su krila nešto proširena. Usne su pune i deblje nego kod Evropida, ali nisu izbunrene. Kosa je, naprotiv, tipično crnačka, gusta i kudrava. Brada i brkovi slabo se očituju. Put je smeđa, s primjesom crvenkastog tona. Ima svjetlijih i tamnijih nijansa.
Etiopidi su temeljna rasa u Abesiniji i Somaliji. Na istoku su jače Pomiješani s Orijentalidima (Arapima), a prema jugu su čistiji. Plemena Amhara, Danakil, Kafičo, Gurage, Gala, Somal, Hima, Masai Pretežno su etiopidne rase.
3. Koisanidna skupina.
Naziv ove skupine složen je po prijedlogu L. Schultzea (1907) od riječi Koi, kako sebe nazivaju Hotentoti, i San, njihove oznake za Bušmane. Niski stas, plosnato lice, nabor na gornjoj vjeđi, steatopigija, posebni oblici spolovila, žućkasta put, uz to fil-fil-kosa, crnačke usne ovu skupinu oštro dijele od svih drugih rasa, tako da je najispravnije dodijeliti joj posebno mjesto. Svakako se radi o rasnoj skupini, koja se valjda rano odijelila od zajedničkog debla Mongolida i Crnaca.
Sanidi (tab. II. sl. 3).
Prosječne mjere: visina 157,7 cm, indeks glave 75,0. Tijelo Bušmana ima uglate forme, tanke su ruke razmjerno kratke, šaka i stopalo uski i mali. Osovine stopala otkrenute su jako jedna od druge, tako se Sanidi mogu lako prepoznati već po tragovima. Donji kraj kralježnice tako je ukrivljen prema straga, da krstača ima gotovo vodoravan položaj. Uglate lične kosti jako iskaču, korijen nosa leži u istoj ravnini s očima, gornja vjeđa ima nabor, koji se spušta preko njenog ruba često prelazeći u pravi mongolski nabor. Usne vire naprijed poput rila, ali nisu izbubrene kao kod Crnaca. Uho je maleno, bez jagodice i koso smješteno. Put je blijedo žućkasto-smeđa ili crvenkastožuta, kao lišće na jesen, dakle prilično svijetla. Potkožne masti gotovo nema, osim na sjedalu, gdje obično stvara prominentan jastuk. Kosa je crna, najgušće kudrava, i stvara male smotane pupoljke. Inače je tijelo bez terminalnih dlaka, brade i brkova. Sanidi živu u suhim stepama sjeverozapadnog dijela kapske zemlje i dalje prema sjeveru.
Koidi (tab. II. sl. 4).
Prosječne mjere: visina 157,9 cm, indeks glave 73,4. Ova rasa, koju su holandski kolonisti nazvali Hotentotima, što znači holandski mucavci, manje je primitivna od Sanida. Stas je nešto viši, glava duža, lice više, sa šiljastom bradom. Otvor vjeđa još je uži nego kod Sanida, a usne su još jače ispupčene, poput rila. Boja kože, oblik i boja kose isti su kao i kod Sanida. Područje Hotentota je istočni dio kapske zemlje i prijašnja njemačka jugozapadna Afrika.
Danas ima u tim područjima već vrlo mnogo različnih mješanaca između Koisanida i drugih rasa, napose bijelaca i Kafrida; naročito su poznati već gore spomenuti križanci u Rehobotu, potomci holandskih useljenika i Hotentotkinja.
4. Mongolidna skupina.
Najznačajnije je obilježje svih Mongolida njihova žućkasta put, koja, doduše, može biti gotovo bijela ili i smeđa, ali uvijek sa žućkastim tonom, zatim glatka, opora, crna ili crno smeđa kosa, a terminalne dlake slabo su razvijene. Kod proporcija tijela upada u oči razmjerno velika dužina trupa. Lične su kosti pomaknute jako naprijed, da nabubruju lice, koje je radi toga, kao i radi udubljenog korijena nosa, plosnato. Otvor vjeđa je uzak i nešto koso smješten, tako da izvanji kraj leži više nego nutarnji. Uz to je kod tipičnih Mongola gornja vjeđa opremljena mongolskim naborom. Mongolidi su bili donedavna brojčano najveća skupina ljudskog roda, te su sačinjavali više od polovice stanovništva čitavoga svijeta. Naziv »Mongoli« potječe od kineske riječi mong-ku, »hrabri«, jer su srednjo-azijski Mongoli svojim upadima i svojom borbenošću zadavali strah svim okolnim narodima.
I kod Mongolida ima morfološki primitivnih tipova s infantilnim obilježjima. Ovi su kao i Paleonegridi obično potisnuti u šumovite i brdovite predjele, ali ima i progresivnih, ekspanzivnih, nizinskih tipova. Prve nalazimo u južnim predjelima istočne Azije i na otocima, dok su drugi prošireni diljem srednjeg i sjevernog pojasa Azije, a došli su i daleko u Evropu, gdje su se njihovi krajnji odvojci pomiješali s Evropidima sve do središnje Evrope.
Paleomongolidi (tab. II. sl. 5—7).
Prosječne mjere: visina 158—162 cm, indeks glave 79—83. Ova je rasa temelj onog pučanstva, koje se obično zove malajsko. Stas je nizak, kod tipičnih predstavnika u južnoj Kini zdepast, dok je na otocima tijelo pravilnije, dapače vitko. Lice je široko, nisko, gotovo kvadratično, kod žena zaokruženo. Nos je plosnat, ali ravan s udubljenim korijenom. Donja čeljust ima slabo naznačenu bradu. Usne su mesnate i naprijed povučene, poduprte lakom alveolarnom prognatijom. Mongolski je nabor na vjeđi naznačen, ali ne seže do nutarnjeg kuta oka. Put je svijetlo smeđa sa žućkastim tonom. Prema ostalim Mongolima pokazuju Paleomongolidi infantilne crte, te su svakako u morfološkom pogledu najprimitivniji mongolski tip. Paleomongolidi zapremaju cijelu južnu Kinu sve do Tibeta, Indokinu, Siam i Burmu, a uz to sve Sundske otoke, gdje im je put nešto tamnija. Na kontinentu se može razlikovati progresivan tip u Burmi; tu podrasu zove Eickstedt Palaungidi. Primitivniji su tipovi u Siamu i dalje prema istoku i sjeveru, šanidna podrasa. Na otocima je Sarasin razlikovao primitivnije Protomalajce u nutrini i progresivnije Deuteromalajce u obalnim predjelima. Prvi su mnogo miješani s Vedidima i Melanidima, te su se svakako ranije uselili nego profinjeni obalni stanovnici, koji imadu polinezidne i indidne primjese. I Eickstedt prihvaća tu podjelu i govori o Protomalajidima i Deuteromalajidima. Paleomongolidi su i znatna komponenta Japanaca.
Tungidi (tab. II. sl. 9 i 10).
Prosječne mjere: visina 162—164 cm, indeks glave 83,5—88,8. Kao što su Sudanidi najtipičniji Crnci, tako se u Tungidima najjače ističu mongolidna obilježja. Stas je zdepast, ruke i noge razmjerno kratke. Lice plosnato, tako da se obrazi nalaze u istoj ravnini s niskim čelom i korijenom nosa. Otvor vjeđa stoji koso, a rub gornje vjeđe pokriven je od mongolskog nabora, koji se uz nos nastavlja sve do donje vjeđe. Usne su nešto punije od evropidnih, bradni šiljak donje čeljusti dobro je razvijen. Kosa je opora, glatka, tamnosmeđe boje. Obrve, brkovi i brada vrlo su rijetki, a tijelo je gotovo bez dlaka. Put je blijedo žuta, bez crvenila. Područje Tungida seže od pustinje Gobi do sjeverne sibirske obale, gdje su pomiješani sa Sibiridima; zapremaju veći dio Mandžurije, a razasuti su i po zapadnoj Sibiriji sve do Volge i Kavkaza. Tunguzi, Mandžui, Giljaki, Burjati, Kalmiki, Ojrati, Tumiti, Kešikti i dr. pretežno su tungidne rase.
Sinidi (tab. II. sl. 11 i 12).
Prosječne mjere: visina 164—168 cm, indeks glave 72—79. Kod Sinida nalazimo sva tungidna obilježja ublažena. Stas je viši i vitak, glava je mesocefalna, dapače dolihocefalna. Nos je uži i nešto povišen, tako da lice nije plosnato. Brada na čeljusti ne iskače tako jako. Često postoji alveolarna prognatija. Otvor je vjeđa širi, ali mongolski nabor obično pokriva nutarnji kut oka. Lice koleba od okruglog do duguljastog, dapače uskog oblika. Uslijed ovog ublaženja mongolidnih obilježja sinidno se lice nešto približava evropidnom tipu. Put je jako svijetla, svijetlo smeđe žuta ili bjelkasto smeđa. Crvenilo je obraza ipak rijetko. Kosa je tipično mongolidna. Sinidi sačinjavaju glavni dio pučanstva središnje Kine, najglavnija su komponenta Koreanaca, a u Japanu su već od davnine pomiješani s Paleomongolidima i Ainuidima. Kod Japanaca postoje dva tipa, finiji Okajama-tip (tab. II. sl. 8) s uskim licem i uskim ravnim nosom, u kojem se ističe sinidna komponenta i grublji Satsuma-tip (tab. II. sl. 7) sa širokim plosnatim licem, gdje prevladava paleomongolidna komponenta.
Sibiridi (tab. II. sl. 13).
Prosječne mjere: visina 156—160 cm, indeks glave 78,5—79,3. Deniker je nazvao ovu rasu ugrijskom, često ih zovu i Paleoazijate. Kao kod Sinida i tu su mongolidna obilježja ublažena. Tijelo je krepko, zbijeno, s dugačkim trupom, ramena široka i čvrsta, silno su otporni protiv studeni, dakle izvrsno prilagođeni na sibirsku klimu. Lice nikako nije plosnato, samo što veliki razmak očiju kvari inače evropidne crte. Nos je obično dosta visok, dapače katkada i kukast. Boja je šarenice svijetlo smeđa, na zapadu ima do 36% modrih i sivih očiju. Kosa je mekša od tungidne, smeđa, često s crvenkastim tonom. Boja je puti svijetlo žućkastosmeđa, na obrazima crvenkasta.
Sibiride nalazimo najjače zastupane u sjeverozapadnoj Sibiriji dalje prema istoku miješaju se s Tungidima, a na krajnjem istoku na Kamčatki, opet su čistiji. Na zapadu su već od davnine miješani s Evropidima, a na istoku s Eskimidima i Indijanidima. Voguli, Ostjaci Keto, Kačini, Kajbali, Kišili, Beltiri na zapadu, a na istoku Čukči i Kamčadali pretežno su sibiridne rase.
Turanidi (tab. II. sl. 14).
Prosječne mjere: visina 166,8 cm, indeks glave 83,6—84,8. Deniker naziva ovu rasu turko-tatarskom. Kao što su Etiopidi očito stara mješavina Negrida i Evropida, tako su Turanidi mongolidno-evropidna smjesa. Montandon i Keith smatraju ih nekim prijelazom između Mongolida i Evropida, a Eickstedt ih, dapače, uvršćuje među Evropide, ali tko pozna tatarski tip, teško će se bezuslovno priključiti tom shvaćanju. Istina je, da kod Turanida nalazimo svih prijelaza od Tungidima sličnih tipova do tipova s mnogo evropidnih crta. Stas je srednje visok, tijelo pravilno, ali naginje ugojenosti. Lične kosti iskaču. Nos je ravan i uzak, ali sa širokim krilima, kakva često opažamo kod Rusa s nešto turanidne primjese, pa i kod nas i u ostaloj srednjoj Evropi. Usne su uske, evropidne. Otvor je vjeđa uzak, što daje licu neki oštar izraz. Mongolskog nabora nema, ali su vjeđe zategnute prema vani na mongolski način. Put je svijetlo smeđa i pokazuje lako crvenilo, ma da se često ističe i nešto žućkasti ton. Kosa je smeđa i lako valovita, nema, dakle, ništa mongolidno.
Turanidi se protežu od Pamira (Pamiridi) preko Turkestana i južne Rusije (Aralidi), do u srce Evrope, sve više pomiješani s Evropidima. Sačinjavaju mongolidnu komponentu u Evropi, kamo su tokom srednjeg vijeka često prodirali kao Avari, Madžari i Tatari. I stari su Osmani bili Turanidi, ali od toga nije ostalo kod današnjih Turaka gotovo ništa, već su od starih Osmana sačuvali samo ime i jezik, a potpuno je preuzela maha u svoje doba od Osmana podjarmljena anatolidna (armenidna) rasa, kako biva svagdje, gdje tanak osvajalački sloj prevlada nekim brojno jakim narodom, čija neiscrpna loza ostane i proguta osvajače.
5. Eskimidna skupina.
Po završetku zadnjeg ledenog doba Mongolima je bio otvoren put preko Beringova tjesnaca, pa je došlo do invazije u Ameriku, a možda se to zbivalo i ranije, u zadnje interglacijalno i glacijalno doba. Na sjeveru Azije tada je bilo i Evropida, što nam posvjedočuju evropidni Ainuidi, koje nalazimo na krajnjem istoku Azije kao zadnji ostatak Evropida. Mongolidi, koji su prodirali u Ameriku, nisu, dakle, bili čisti Tungidi, nego miješani s evropidnim elementima; stoga se ne smijemo čuditi, što se kod američkih urođenika, Eskimida i Indijanida, javljaju mnoga evropidna obilježja. Eskimidna je skupina zastupana jednom jedinom rasom, ali je ta obilježena tolikim posebnim svojstvima, da za služuje položaj naročite skupine.
Eskimidi (tab. II. sl. 15 i 16).
Prosječne mjere: visina 157,4—164,8 cm, indeks glave 77,6—80,7. Tijelo je nisko i zdepasto; visina je tijela kod Eskima na zapadu veća, a prema istoku sve manja. Trup je dugačak, udovi, napose bedra, kratki su. Ramena su krepka i široka. Mišići su čvrsti, ali golijeni tanke. Šaka su i stopalo veoma kratki i maleni. Glava je prema niskom tijelu vrlo velika, često nezgrapna, čelo nagnuto. Preko tjemena proteže se uzdužna uzvisina. Lice je plosnato i veoma široko, najšire od lica svih ljudskih rasa, jer su lične kosti široke, i stražnji uglovi donje čeljusti jako razmaknuti. Nos je ravan, s visokim hrptom, prilično uzak (indeks 68—72). Usne su evropidne, samo se često javlja lagana alveolarna prognatija, koja usne nešto izdiže. Uho je veliko i usko. Put je smeđe žuta ili crvenkasto smeđa. Obrazi su crvenkasti, kao kod Evropida. Šarenica je smeđa rijetko (1%) modra. Kosa je glatka ili lako valovita, crna. Obrve su obično jako razvijene. Dlakavost je slaba. Brada je slaba i rijetka, katkada crvenkasto smeđa. Ma da su Eskimidi prošireni velikim područjem, u somatskim su svojstvima prilično jedinstveni.
Eskimidi sežu od Alaske (i nešto naselja na azijskoj strani Beringova tijesna) do istočne Grönlandije, ali su njihova obitavališta rijetka, tako da ih ima ukupno kojih 40.000.
6. Indijanidna skupina.
Prema brojnim indicijama prešli su predšasnici Indijanida u vrlo rano kulturno doba iz Azije u Ameriku. Oni tada još nisu poznavali pisma, a nije im bio poznat ni kotač. Svakako se prelaženje iz Azije dešavalo u više valova. Jesu li se zbile prve seobe već za vrijeme diluvija ili tek poslije zadnjeg ledenog doba, u tome su mišljenja podijeljena. Hrdlička, Rivet i Steinmann zalažu se za postglacijalnu seobu, dok Boas, Penck, Birket-Smith, Goddard i dr. drže, da je bilo seoba i u diluviju. Ima u Americi i nekih ostataka kamenog oruđa od Trentona, kojemu i Boule pripisuje diluvijalnu starost. U wisconsinsko ledeno doba, koje u biti odgovara würmskom u Evropi, bilo je u Americi više povlačenja i napredovanja leda, tako da je bilo u onim toplijim fazama moguće prelaziti preko Beringova mora. Valovi kasnijih useljenika potiskivali su ranije useljenike prema jugu, tako da imamo baš u Južnoj Americi očekivati potomke najranijih useljenika. Vjerojatno je, da pojedini valovi useljenika nisu bili po svojoj rasi jednaki i da je to jedan od uzroka rasne pocijepanosti Indijanida. Svakako je moralo dovesti i prilagođivanje na tako raznolike klimatske prilike pojedinih američkih predjela do daljnjeg stvaranja rasnih razlika.
Svim Indijanidima zajednički je žućkasto smeđi ton boje puti, crna glatka kosa i slabo razvijena dlakavost tijela. Oznaka »crvenokošci« nema s prirodnom bojom kože nikakve veze, već potječe odatle, što su neki Indijanci običavali kod pripreme za borbu i u svečanim zgodama mazati lice crvenom bojom. Šarenica oka je smeđa, a otvor vjeđa uzak. Katkada se javlja i mongolski nabor. Nos je prominentan, dapače velik, često orlovski i prilično uzak (indeks 70—76). Kralježnica je malo ukrivljena, tako da su leđa plosnata. Hrd1ička ističe, da je kod svih Indijanida omjer između nadlaktice i podlaktice kao i između bedra i golijeni gotovo isti. Širina je ramena i kukovlja jednaka. U drugim svojstvima, na pr. u visini tijela, indeksu glave, intenzitetu boje puti ima velikih razlika.
Rase Indijanida možemo podijeliti u dvije, izvjesnim razlikama obilježene grupe, južnoameričku i sjeverno-američku. Prva grupa, koja je i najstarija, sadržava najprimitivnije tipove.
a) Južni Indijanidi.
Već je A. d’Orbigny (1839) razlikovao 3 rase u Južnoj Americi: andsko-peruansku, pampasku i brazilsko-guajansku. Ovima treba dodati još rasu, koja živi u brazilskim brdima, a od koje ima i fosilnih ostataka u spiljama Lagoa Santa i onu primitivnu rasu u Ognjenoj zemlji (Tierra del Fuego), koja pokazuje neka obilježja najstarijih fosilnih ostataka iz južne Amerike. Eickstedt upotrebljava za te rase nazive: Lagidi, Fuegidi, Patagonidi, Brazilidi i Andidi.
 tabla III.
tabla III.Lagidi (tab. III. sl. 1).
Prosječne mjere: visina 158,6—161,8 cm, indeks glave 76,8—80,2. U istočnoj Braziliji, u spiljama tamošnjeg vapnenačkog gorja, g. 1840 pronašao je Lund fosilne ostatke čovjeka. Najbogatija je nalazima bila spilja u području Lagoa Santa, sa kojih 30 okosnica. Starost se nije dala točno odrediti, ali svakako su kosti vrlo stare, jer su potpuno okamenjene. Lubanje imadu naročiti tip s uzvišenim tjemenim dijelom kao sljeme na krovu. Te su nalaze nazvali Lagoa-Santa-rasom. Već g. 1876 ustanovio je Lacerda, da ta rasa nije, kako se mislilo, izumrla, već da se fosilne lubanje poklapaju u bitnim potezima s lubanjama današnjih Botokuda, a Rivet (1908) je pronašao, da isto vrijedi i za različna druga plemena onog područja. Radi se, dakle, o rasi, koja je svakako vrlo stara, za to ju je Deniker nazvao paleoameričkom.
Trup Lagida je čvrst i muskulozan, i udovi su čvrsti, a mišići golijeni dobro razvijeni. Lice je primitivno i grubo, u obliku pravokuta, nadočni su lukovi naglašeni. Nos ima široka naduvena krila. Kosa je opora, glatka, crna. Put je svijetlo žutosmeđa, kod novorođenčadi svijetlo ružičasta kao kod Evropida. Područje Lagida su brdoviti predjeli u istočnoj Braziliji. Plemena Karaja, Nahukva, Botokudi, Ges lagidne su rase. Tragova lagidne rase ima primiješanih Patagonidima sve do Ognjene zemlje, gdje se lagidni tipovi opet jasnije ističu, tako napose kod plemena Alakaluf i Jamani, koji su vrlo niskog stasa (154,7 cm).
Fuegidi.
Prosječne mjere: visina 172,9 cm, indeks glave 78,8. Uz brazilsku obalu nađene su u naslagama fosilnih školjaka lubanje, koje pokazuju, kao i vrlo stara — možda diluvijalna — fosilna lubanja, nađena kod Punina u Ekvadoru, australidna obilježja, nadočne nabubrine uz prognatiju i sljemenasto tjeme, kojim se ističe i lubanja iz Lagoa-Sante. Neki potezi tog australidnog fosilnog tipa nalaze se kod nekih sadašnjih plemena u Ognjenoj zemlji, napose kod plemena Ona. Inače pokazuju Ona-Indijanci mnoge crte Patagonida. Osobito su visokog stasa i herkulske konstitucije. Australidne poteze te i s obzirom na kulturni stupanj vrlo primitivne rase možemo samo tako razumjeti, da se radi o najstarijim useljenicima Amerike, koji su došli preko Alaske, kada je živio u susjednoj Aziji još prvobitni australidni sloj.
Patagonidi (tab. III. sl. 2).
Prosječne mjere: visina 161,6—172,9, indeks glave 77,6—78,8. Po fantastičnim pripovijestima španjolskih putnika o patagonskim orijasima već je ova rasa dugo poznata, samo što se kasnije ispostavilo, da nisu viši od prosječnog Evropejca. Patagonidi su rasa lovaca u prostranim Pampasima. Tijelo im je širokih ramena, jako mišićavo, tako da se često javljaju atletske konstitucije. Čelo je nisko, s jakim nadočnim lukovima, otvor vjeđa, koje su zategnute, vrlo je uzak. Uglovi su donje čeljusti razmaknuti, tako da je lice uglato i široko. Obrazi su naduveni. Nos je prilično velik i ravan. Put je obično smeđa. Kosa je glatka i crna. Područje su te rase stepe Pampasa, Chaca i Patagonije sve do Ognjene zemlje. Plemena Baikuru, Čoroti, Čarua, Puelče, Tehuelče pretežno su patagonidne rase. Prema sjeveru su zadnji ogranci Patagonida plemena Čikito u Chacu i Bororo na gornjem Rio dos Montes.
Brazilidi (tab. III. sl. 3).
Prosječne mjere: visina 157,3—160,6 cm, indeks glave 79,1—81,2. Tijelo je čvrsto, muskulozno, ali ni izdaleka ne tako, kao kod Lagida. Ruke su i noge prilično tanke. Lice je ovalno, s ravnim čelom, lične kosti ne iskaču mnogo. Nos je srednje dužine i ravan. Ima mnogo evropidnih crta u licu Brazilida. Koža je žutosmeđe boje kao glina. Kosa je glatka ili valovita, tamno smeđa. Šarenica također tamno smeđa, a bjeločnica ima žućkasti ton. Brazilidi su naseljeni u prašumama uz rijeku Amazonku, u brdovitim predjelima Guajane, gdje se spuštaju do obale i sežu preko svih antilskih otoka, a prema jugu do ušća rijeke La Plata. Glavne su etničke grupe brazilidne rase Karaibi, Aruak i Tupi. U Guajani ima mnogo križanaca između bivših crnačkih (sudanidnih) robova i Brazilida, tako da tvore posebna plemena t. zv. šumskih Crnaca.
Andidi (tab. III. sl. 4).
Prosječne mjere: visina 157,0—158,4 cm, indeks glave 82. Tijelo je nisko i zdepasto, s jako proširenim grudnim košem poput bačve, što je prilagođivanje rijetkom zraku visokoga gorja. Trup je dug, tako da se čini, da su noge, ispoređene s trupom, slabunjave, ma da je njihovo mišičje i na golijenima dobro razvijeno. Vrat je kratak, kao da glava sjedi neposredno na trupu. Čelo je nisko i široko, a tjeme se uzdiže polagano, tako da postizava svoj vrh daleko straga. Lične kosti su naglašene. Oči su prilično razmaknute. Rub gornje vjeđe s trepavicama nastavlja se preko nutarnjeg kuta oka, dok je mongolski nabor rjeđi. Iz plosnatog lica jako iskače visoki nos, ravan ili kukasto ukrivljen. Oštri se nabori povlače od nosnih krila prema kraju usta, kao kod Dinarida. Koža je maslinasto smeđa i prilično tamna, napose kod stanovnika suhih sunčanih visoravni. Kod žena pokazuju obrazi, unatoč tamne boje, jasno crvenilo. Obrve su rijetke. Kosa je gusta, glatka, smeđe crna. Šarenica je tamno smeđa, a bjeločnica žućkasta. Andidi su rašireni diljem lanca visokih Anda, u području nekadašnjeg carstva Inka, sve do južnih čilskih bregova. Andidi se javljaju u tri glavna tipa, koji se podudaraju s etničkim grupama Kečua u Peruu, nižeg stasa, Ajmara u Boliviji, koji su višeg stasa, i borbenih Araukanaca u Čileu, koji su najvišeg stasa (oko 164 cm), s crtama, koje često sjećaju na evropidna obilježja, pogotovo, jer su svjetlije puti.
b) Sjeverni Indijanidi.
U somatskom pogledu pokazuju Indijanidi sjeverne i središnje Amerike više evropidnih crta nego južnoamerički, naročito s obzirom na svjetliju boju kože i oblik nosa kod progresivnih tipova. Mogu se razlikovati četiri skupine: središnji Indijanidi, onda primitivan mesocefalan tip srednjeg stasa na jugozapadnom rubu i Floridi, brahicefalan tip niskog stasa uz ostali dio obale Tihog oceana i progresivan mesocefalan tip vrlo visokog stasa u prerijskom području USA i Kanade. Eickstedt upotrebljava za te rase nazive: Centralidi, Margidi (lat. margo »rub«), Pacifidi i Silvidi (lat. silva »šuma«)«
Centralidi (tab. III. sl. 5).
Prosječne mjere: visina tijela na sjeveru 169—172 cm, u srednjem dijelu 155—159 cm, na jugu 158—166 cm; indeks glave 79—86. Centralidi su ona rasa, koja je proizvela najviše američke samorodne kulture, Azteki i Maja su centralidnog podrijetla. Tijelo pokazuje donekle evropidne razmjere, jer trup nije velike dužine. Lice je kod sjevernih grupa razmjerno usko, dok se visoko čelo prema tjemenu suzuje, što je za Centralide osobito značajno. Budući da su uglovi čeljusti dosta razmaknuti, ima lice pravokutan oblik. Otvor je vjeđa širi nego inače kod Indijanida. Mongolskog nabora gotovo nema. Šarenica je tamno smeđa; glatka, crno smeđa kosa naraste vrlo duga i lako valovita. Za razliku od većine drugih Indijanida kosa Centralida vrlo rano posijedi. Dlakavost je nepotpuna, i na bradi. Boja puti je varijabilna, obično je srednje tamno smeđa. Vrlo se često javlja albinizam, najčešće kod plemena San-Blas na Darjenskom tjesnacu. R. O. Marsh je g. 1924 ponovno otkrio te bijele Indijance, pa je poslana posebna ekspedicija, da ih proučava, jer se držalo, da se radi o nekim evropidnim elementima, ali se ispostavilo, da je to albinotička pojava, koja se je radi endogamije veoma umnožila.
Centralidi su prošireni od Kolumbije i Venezuele preko Panamskog tjesnaca i istočnog Mehika do donjeg Misisipija i atlantske obale sjeverno od Floride. Pojedini eksklavi dostižu Mičigensko jezero. Čičimeki, Azteki, Mixteki, Zapoteki, Maskoki uglavnom su Centralidi. I danas se ovi potomci tvoraca visokih kultura, unatoč zapostavljanju od bijelaca, ističu svojom odrešitošću i razboritošću. Centralidi su danas mnogo miješani s bijelcima, napose u Mehiku i u državama tjesnaca, to su Mestici ili Ladinos; ima mnogo i indijansko-crnačkih križanaca, nazvanih Zambo ili Kaboklo.
Margidi (tab. III. sl. 6).
Prosječne mjere: visina 163—170 cm, indeks glave 78,5—79,5. Sjeverniji tip ove rase nižeg je koščatog stasa, dok je južni višeg stasa. Čelo je plitko, tjeme pokazuje uzdužnu uzvisinu kao kod Eskima. Malo je oko natkriljeno izrazitim nadočnim lukovima. Korijen konkavnog nosa leži duboko. Vjeđe su bez mongolskog nabora i nisu napete. Lice ima dosta mekane crte. Boja je kože tamno smeđa s crvenkastim tonom. Kosa je glatka, crna, dlakavost slaba. Kalifornija i visoravni između obadva lanca Kordiljere sve do tjesnaca obitavališta su ove rase. Hrd1ička je po lubanjama pratio taj tip sve do središnjeg Mehika, gdje je ova rasa sačinjavala prastanovništvo prije nego su došli Azteki. Pojedini ostatci te rase raštrkani su i preko tjesnaca, sve do južne Amerike, gdje ih ima u Britskoj Kolumbiji. Šošoni, Juma, Otomi plemena su pretežno margidne rase.
Pacifidi (tab. III. sl. 7).
Prosječne mjere: visina tijela 167,3—169.7 cm. indeks glave 84,2—88,8.
Tijelo je masivno građeno, trup kratak, a ruke duge. Šaka i stopalo su veliki i široki. Čelo je visoko, nos vrlo sličan nordidnome nosu, samo što je njegov hrbat nešto širi, a tako i donji dio. Kako ni lice nije plosnato, njegov je izraz sličan evropidnom licu. Usta su uska, a prednji dio donje čeljusti masivan, kako to često vidimo i kod evropskih naseljenika u Americi. Nazolabijalni nabori naglašeni su. Mongolskog nabora na vjeđi nema. Boja kože je jako svijetla, svijetlo smeđa ili žućkasta, tako da neki izvještaj prvih španjolskih osvajača kaže: »tan blancos que los mejores Españoles« (bijeli, kao najbolji Španjolci). Kosa je valovita i tamno smeđe boje, češće i svjetlija. Područje su Pacifida šumoviti predjeli zapadne Kanade uz otoke i susjedna Alaska, a prema jugu sežu do sjeverne Kalifornije. Plemena Tlinkit, Haida, Cimšijan, Kvakjutl, Nutka, Heilcuk gotovo su čisto pacifidne rase.
Silvidi ili Šumski Indijanci (tab. III. sl. 8).
Prosječne mjere: visina 170—175 cm, indeks glave 79—81. Silvidnoj rasi pripadaju oni »pravi« Indijanci, koji su nam poznati iz omladinske književnosti po svojoj hrabrosti i svom dostojanstvu. Algonkini, Delavari, Huroni, Tuskarore, Siu, Komanči, Odžibve i t. d. pretežno su Silvidi.
Tijelo se Silvida razlikuje od evropidnih proporcija po tome, što je trup razmjerno kratak, ali širok; trup se čini zdepast, a udovi su dugački. Napose široka ramena daju tijelu izražaj čvrstoće i jakosti, pogotovu, jer je grudni koš prostran. Glava je razmjerno velika, čelo široko i nisko, lične kosti dosta široke, mongolidne, otvor je vjeđa uzak, ali stoji vodoravno, mongolskog nabora nema. Usta su široka, s uskim usnama. Nos je visok i velik, obično orlovski ukrivljen, vrlo sličan dinaridnom nosu, oštri nazolabijalni nabori protežu se od nosa prema krajevima usana, pa tako lice dobiva odrješit izražaj. Kosa je glatka, crna, dlakavost pak neznatna, a gotovo nema ni brkova ni brade. Šarenica je tamno smeđa, a koža svijetlo smeđa, slična kao kod pigmentiranih Evropida, na suncu dobivaju lak crvenkasti ton, jer prosijavaju krvne žilice. Tako je i koža na obrazima djevojaka crvenkasta.
Svih Silvida ima danas u Kanadi još kojih 100.000, a u prerijama, u rezervacijama USA kojih 80.000, žalosni ostatci nekad tako moćne i ponosne rase.
7. Evropidna skupina.
Svijetla put, na obrazima ružičasta, bujna, tanka, valovita kosa, priličan razvitak terminalnih dlaka, obrazi u poprečnom smjeru zaokruženi, najšire mjesto jagodica natrag pomaknuto, ravno visoko čelo, uzak nos s uskim krilima i uzvišenim korijenom (među očima), ravnozube čeljusti, donja s naprijed izbočenim bradnim šiljkom, proporcije tijela, koje donekle odgovaraju umjetničkom kanonu; to bi bila obilježja, koja jasno ograničuju evropidnu skupinu. Ako je zovemo evropidnom, to još ne znači, da je nalazimo samo u Evropi. Jednake tipove, bilo u kompaktnoj masi, bilo razasute i miješane, naći ćemo kao autohtone u cijeloj sjevernoj Africi i dalje do Burme, pa opet na južnom dijelu otoka Sahalina i na otoku Jeso te dalje preko polinezijskih otoka sve do Nove Zelandije. Odakle potječu ti ljudi? Da to razumijemo, treba se obazreti na prilike u zadnje ledeno doba. Neprohodne zapreke, ledenjaci i ledene stepe dijelile su tada središnji dio Azije od jugozapadnih, južnih i jugoistočnih predjela. Vrlo uska staza, negdje u području današnjeg grada Kazana, vodila je u onaj dio Evrope, koji nije bio pod ledom, ali je zato bio većim dijelom hladna stepa, koja nije pružala mnogo mogućnosti za opstanak. Neandertalac je tada ondje gospodovao. Lako je bilo doći u Evropu od juga i jugoistoka. Možda je aurignacski čovjek došao tim putem, pa njegova obilježja imaju neke australidne crte. Jednako je očito, da je došao od juga i grimaldijski čovjek.
Ali i Cro-Magnonac već je bio izabrao negostoljubive predjele tadašnje Evrope svojim obitavalištem. On je bio očiti Evropid. Evropidi na Sahalinu i Jesu nipošto nisu mogli doći onamo preko Kine i Mandžurije, jer bi se prije utopili u ondješnjem pučanstvu, nego su mogli doći samo iz sjeverne Azije, kad je iščezla početkom aluvija ledena barijera na istoku Sibirije. Sve nas to upućuje na to, da Evropidi potječu iz središnje Azije. Cromagnonci su bili njihova prethodnica. Kada se počeo led otapati, nastupile su u središnjoj Aziji povoljnije životne prilike, pučanstvo se počelo razmnažati, njegova je ekspanzija bivala sve veća. Na jugoistoku naišla je na otpor Mongolida, te su se probile na istoku samo manje skupine do Sahalina, i to valjda, sudeći po primitivnom obilježju Ainuida, vrlo rano. Na zapadu, u onoj tada nenastanjenoj Evropi, nije bilo otpora, pa su počeli dolaziti bijelci u migracionim valovima u novu, od leda oslobođenu zemlju. I iranska je barijera popustila, pa su druge iseljeničke struje upale u Indiju, Mezopotamiju, odatle u Malu Aziju i prema jugozapadu u Siriju i Arabiju te dalje u Afriku, u Španjolsku i ostala sredozemna područja.
Bijelci tih različnih migracionih valova bili su svakako već pocijepani na rase, jer je u središnjoj Aziji bilo toplijih i hladnijih predjela, više ili manje odijeljenih, koji su pogodovali stvaranju rasa. Poslije Ćro-Magnonaca najprije su doprli brahicefalni ljudi, kako pokazuju fosilne brahicefalne lubanje iz tog ranog doba, a za njima preko male Azije drugi brahicefalan val, iza kojega su slijedili jedan dolihocefalan val na sjeveru, kroz južnu Rusiju, a drugi na jugu preko Mezopotamije u Afriku i Mediteran. Valjda je počela tada već ekspanzija Mongolida, koji su od istoka dopirali u središnju Aziju i potiskivali bijelce prema zapadu. Kao zadnji došao je u Evropu opet val brahicefalnih bijelaca već u povijesno doba, a za njima su nagrnuli u Evropu Mongolidi, kao Huni, Avari, Madžari, Pečenezi, Kumani i Tatari, jer je i njima postala središnja Azija tijesna. To je najispravnija slika, u koju se mogu složiti geološke činjenice, podatci fosilnih i pretpovijesnih nalaza, kao i povijesne tradicije.
Evropidi, pogotovu njihovi primitivniji tipovi, imadu mnogo crta zajedničkih s Australidima, tako da je vjerojatno, da su njihovi predšasnici bili australidni tipovi, koji su u srednjem diluviju doprli u središnju Aziju te se tu dalje diferencirali u Evropide. Aurignacski tip bio je možda početni stadij.
Do sredine prošlog stoljeća Evropidi, nazvani kavkaskom ili leukodermnom rasom, bili su smatrani većinom kao jedinstvena rasa, ma da je već Desmoulins (1825) razlikovao skitsku, kavkasku, indijsku, atlantsku i kurilsku (Ainu) »vrstu«, a Bory de St. Vincent (1827) jafetitsku, arapsku i indijsku »vrstu«. Tek mjerenja A. Retziusa pokazala su, da ima dolihocefalnih i brahicefalnih bijelaca, pa da ima i u Evropi između njih rasnih razlika. W. Z. Ripley (1900) razlikovao je u Evropi tri rase: teutonsku, alpsku i mediteransku. Prva bi odgovarala Denikerovoj nordijskoj rasi, mediteranska sadašnjem shvaćanju, a u alpsku rasu udružio je sve brahicefalne tipove. Već je F. v. Luschan (1889) istaknuo, da treba razlikovati poseban armenoidni ili hetitski tip, koji danas obično zovu prednjoazijskom ili armenidnom rasom. S obzirom na načelo rasnog nazivlja, da se za rase nikad ne upotrebljavaju etnička ili lingvistička imena, mi ćemo ovu rasu zvati anatolidnom (Anatolija »Prednja Azija«). Napokon je Deniker (1900) dao svoju podjelu rasa, koja je u bitnim crtama općenito prihvaćena. Već je La Pérouse (1797) ispravno naglasio, da su Ainu bijelci, ali je to zaboravljeno, dok nije Montadon (1927), najbolji poznavalac tog tipa, naišao na opće priznanje svojom tvrdnjom, da se tu radi o Evropidima. Da i Polinezijci pripadaju Evropidima, istaknuo je E. Fischer (1931) u svom rodoslovlju evropidnih rasa; Eickstedt zauzima isto stajalište, pa somatička su svojstva Polinezijaca u svim bitnim smjernicama evropidna.
a) Oceanski Evropidi.
Ainuidi (tab. III. sl. 9 i 10).
Prosječne mjere: visina 157,2—158,0 cm, indeks glave 76,8—77,8. Trup je širok, masivan. Lice je nisko. Čelo iskače s nadočnim lukovima naprijed, te oči leže duboko. Donja je čeljust jako krepka i sprijeda plosnata, jer bradni šiljak nije naglašen. Često postoji, za razliku od svih ostalih Evropida, lagana alveolarna prognatija. Nos je srednje visine, prilično širok, često s konkavnim hrptom, da su otvori sprijeda vidljivi, napose kod žena. Koža je blijedo bijela, često ružičasta. Kosa je bujna, gusta, lako kovrčasta, smeđe boje. Kod muškaraca ima dugih brada; prsa i leđa, isto i bedra pokriveni su gustom dlakom. Od svih rasa Ainu imaju najrazvijenije vlasi i dlake, pa se po tome osobito oštro razlikuju od okolnih Mongolida, koji su gotovo bez dlaka i imaju rijetku bradu. Kako su brkovi i brada kod Ainu u cijeni, pokazuje navika žena, da sebi tatauiraju brkove, kako ne bi imale tako praznu usnu. Oblik glave pokazuje, da su Ainuidi stara primitivna rasa, koja se donekle priključuje na australsku skupinu, najprvobitniji bijelci. Većina je lako miješana s Mongolidima, što pokazuju široki obrazi s naglašenim jagodicama. Katkada se javlja i mongolski nabor. Ali ima i pojedinaca bez ikakvog mongolidnog obilježja.
Ainuidi živu na najsjevernijem japanskom otoku Jeso, na južnom kraju Sahalina i na Kurilskim otocima, koji se odatle protežu do Kamčatke. Prije je njihovo područje bilo veće. U srednjem vijeku postojale su na glavnom japanskom otoku Hondo posebne Ainuske kneževine. Poslije su se ti Ainu utopili u japanskom pučanstvu, koje često pokazuje pojedine ainuidne crte.
Polinezidi (tab. III. sl. 11 i 12).
Prosječne mjere: visina 173,7—171,1 cm, indeks glave 77,7—84,0. Da ovu rasu navodimo poslije Ainuida, to ne znači, da su Polinezidi po svom podrijetlu u nekoj bližoj vezi s Ainuidima, već jedino to, da i oni sačinjavaju prema ostalim Evropidima neku zasebnu skupinu, ali više s obzirom na neevropidne rasne elemente, koje nalazimo u manjoj ili većoj mjeri primiješane Polinezidima. Evropidna komponenta je dapače bliska Mediteranidima. Polinezijci su kao drski i odrešiti pomorci došli u svoja sadašnja obitavališta dosta kasno, u toku zadnjih dvaju tisućgodišta. Najkasnije, oko 1400 g., zaposjeli su Novu Zelandiju i podjarmili melanezidne prasjedioce. Predaja Polinezijaca govori o tome, da su njihovi predšasnici došli iz zemlje, gdje su stabla samo pol godine zelena i gdje se moglo hodati po vodi (led!), uz to govori njihova tradicija o plavokosim ljudima.
Tijelo je Polinezida vitko, muskulozno, s evropidnim proporcijama. Mišići golijeni osobito su jako razvijeni. Žene su znatno manje od muškaraca, njihovo tijelo naginje bucmastom obliku, gležnjevi su nešto deblji nego kod Evropejaca. Lice je ovalno, čelo visoko i široko, nos prilično uzak, ravan, s visokim korijenom i nešto mesnatim krilima. Usne su uske, tipično evropidne. Otvor je vjeđa širok, samo što se kod 50% svih Polinezijaca javljaju tragovi mongolskog nabora, a kod 5% je nabor jače izražen, posljedica toga, što imaju primiješane nešto paleomongolidne krvi. Boja je kože svijetlo smeđa s crvenkastim tonom, katkada je i sasvim svijetla, a ima svih prijelaza do maslinasto smeđe boje. Kosa je bujna, valovita do lako kovrčaste, obično tamno smeđa. Obrve, brada i brkovi dobro su razvijeni, dok je inače dlakavost tijela slaba. Kod pojedinaca javlja se i plava kosa, napose kod djece. Šarenica je smeđa, pojedinačno siva ili svijetlo zelena.
Već Cook i ostali moreplovci, koji su sredinom 18. st. otkrili oceanske otoke, ostali su začuđeni, da su ondje našli ljude, gotovo sebi jednake, a ne neke divljake. Napose hvale svi, koji su upoznali Polinezijce, njihovu otmjenu suzdržljivost, veselu i otvorenu ćud, te prijaznost i srdačnost. Polinezijci vrijede kao vrlo lijepi ljudi, napose ističu se žene velikom ljupkošću, a ima ih mnogo, koje se mogu uvrstiti među najljepše, prema evropskom estetskom mjerilu. Na žalost, djeluje i na taj svijet dodir s Evropejcima porazno. Sifilis, alkohol i druge slične »blagodati« naše civilizacije i gramzljivost kolonista doveli su i ovu žilavu i inteligentnu rasu na rub propasti.
Područje Polinezida su otoci Tihog Oceana od Havaja do Nove Zelandije. Najčišća se rasa nalazi na Samoanskim otocima, ali i tu imaju primiješane nešto paleomongolidne i melanezidne krvi, drugdje je ta primjesa jača. Na Havajskim otocima imaju Polinezidi nešto grublje crte lica, jednako na Tongi, glava je kod tog tipa nešto okruglija. Melanezidna primjesa najjača je na otocima, koji su bliže melanezidnim područjima, tako napose na Fidži-otocima. Stanovnici Mikronezijskih otoka imadu pored melanezidne i prilično mnogo paleomongolidne primjese, pa im je lice plosnato, a put bakreno smeđa. Stoga predstavljaju poseban jasno ograničen tip; Eickstedt govori o mikronezidnoj podrasi (tab. III. sl. 13).
U novije doba ima mnogo križanaca Polinezida s evropskim bijelcima, a na Havajima i s Kinezima.
b) Azijski Evropidi.
Indidi (tab. III. sl. 14 i 15).
Prosječne mjere: visina 159,8—163,3 cm, na sjeveru 171,9—172,1 cm; indeks glave 72,2—75,9. Oznaka Indidi poklapa se s Denikerovom indo-afganskom rasom. Tijelo je Indida vitko i lijepo proporcionirano. Podlaktice i golijeni su tanke, što povećava gracilnost tijela. Čelo je uspravno, usko i stoji naprijed iznad jakog korijena nosa. Nos je uzak, ravan. Usne su nešto deblje nego kod evropskih rasa. Lice je usko, ovalno, kosa je valovita, crno smeđa. Put je svijetlo do srednje-tamno smeđa. Uglavnom se daju razlikovati dva tipa: sjeverni Indidi iz Pandžaba, visokog stasa i svjetlije puti, i Indidi iz Hindustana i Dekana, koji su manji po stasu, gracilniji, ali i nešto tamniji.
Područje Indida obaseže srednji i istočni Afganistan, Beludžistan, sjeverni dio obale Indije kao i središnji Dekan. U različnim predjelima tog područja ima mnogo lokalnih tipova te rase. Indidna je rasa bitna komponenta cijelog niza indijskih naroda i južnih grupa, ne samo onih, koji govore arijske jezike. Ma da propisi kasta donekle zaštićuju Indide od miješanja s drugim indijskim rasama, ipak je iz nižih slojeva postepeno doprla vedidna i melanidna krv, sve do najviših kasta.
Indidne rase bili su prvobitno Cigani; još danas je kod njih indidna komponenta vrlo jaka, ma da su kod svojih seoba primili i rasne komponente svih zemalja, kroz koje su prolazili.
Orijentalidi (tab. III. sl. 16).
Prosječne mjere: visina 162,8—169,8 cm, indeks glave 76—78,8. Kod stanovnika pustinja stas je vitak, žilav, ali kod prijelaza na život u gradovima javlja se nagnuće gojaznosti, napose se to opaža kod žena. Lice je ovalno, srednji dio lica ima neku mekoću, koja se očituje na debelim usnama. Nos je vrlo uzak s visokim hrptom, ravan i lagano ukrivljen. Otvor vjeđa ima oblik badema, u nutarnjem kutu zaokružen a prema vani sužen. Koža je smeđa u različnim nijansama. Šarenica je tamno smeđa. Kosa je crno smeđa i kovrčava.
Ova je rasa prije često nazivana semitskom u vezi s tim, što mnogi predstavnici te rase govore semitske jezike. Područje je orijentalidne rase Perzija, Mezopotamija, Sirija, Arabija i sjeverna Afrika. Najtipičniji su predstavnici te rase Arapi. U većem dijelu toga područja Orijentalidi su već pomiješani s anatolidnim (armenidnim) i turanidnim elementima, a u Africi s Negridima. Abesinsko pučanstvo, napose na istoku, ima također orijentalidne primjese. Orijentalidi su bitna rasna komponenta Židova, napose sefardijskih (španjolskih), dok aškenazijski Židovi imaju pored orijentalidne napose anatolidnu komponentu uz primjese gotovo svih evropskih rasa. Orijentalidi su i Parsi u Indiji, koji su se onamo doselili iz Arabije u 7. st. i radi svoje vjere, koja im zabranjuje miješanje s inovjercima, sve do danas sačuvali svoju rasu.
 tabla IV.
tabla IV.Anatolidi (Armenidi) (tab. IV. sl. 1).
Prosječne mjere: visina 165,5—167,2 cm, indeks glave 84,2—85,2. Ova brahicefalna južnjačka rasa poznata je već s hetitskih spomenika, kako je to pokazao Luschan. Dakle već u drugom tisućljeću pr. Kr. ova se rasa istakla kao tvorac visokih kultura. Stas je nešto zdepast, jer su noge razmjerno kratke. Tijelo naginje gojnosti, napose u kasnijim godinama. Već ranije ističe se gojnost kod žena, pa to odgovara istočnjačkom idealu ljepote. Najtipičnije obilježje Anatolida je veliki ukrivljeni nos, najveći nos, što ga ima neka ljudska rasa. Krajnji dio nosa obično je još kukasto ukrivljen prema dolje. Lice je usko s visokim čelom, usta široka, uho veliko. Valovita je kosa tamno smeđa, isto i šarenica; brada i brkovi dobro su razvijeni.
Područje Anatolida je Mala Azija, kavkaske zemlje, sjeverna Sirija, istočna Perzija, Turci i Armenci pretežno su anatolidne rase. Anatolidna je rasa i bitna rasna komponenta Židova. T. zv. židovski nos anatolidni je nos, kakav nalazimo i kod Turaka, Armenaca i Kavkazaca.
c) Evropski Evropidi.
Dok kod dosele prikazanih rasa ima ipak pojedinih središta, pa i većih područja, gdje dotična rasa čini glavni dio izvjesnog sloja pučanstva, kao na pr. u Indiji, u Evropi ni izdaleka ne postoje više takve prilike, nego su tu rase jako ispremiješane, tako da možemo naći diljem cijele Evrope sličnih tipova, samo što su pojedini tipovi u različnim predjelima brojčano raznoliko zastupani. Samo na krajnjem jugu i krajnjem sjeveru nalazimo donekle neku jedinstvenost. Tek ako promatramo prosječnu podjelu somatskih svojstava, kao visinu tijela, indeks glave, lični indeks, pigmentaciju, ustanovit ćemo: 1. da se u južnim predjelima gomilaju tipovi nižeg stasa, tamnih kompleksija, duge glave, uskog lica; 2. u jugoistočnim tipovi visokog stasa, brahicefalni, tamnih kompleksija, uskog lica; 3. u srednjem pojasu tipovi niskog stasa, okrugle glave, širokog lica, tamnijih kompleksija; 4. dalje prema istoku ljudi sličnog stasa i glave, ali svijetlih kompleksija; 5. u sjevernim predjelima ljudi visokog stasa, jako svijetlih kompleksija, dolihocefalni, uskog lica. Tako dolazimo do pet glavnih rasa u Evropi: Mediteranida, Dinarida, Alpida, Baltida i Nordida. Uz to ima i nekih manjih skupina: na sjeverozapadu i sjeveru Dalide, a kao odvojak Alpida na krajnjem sjeveru Lapide (među Laponcima).
Mediteranidi (tab. IV. sl. 2 i 3).
Prosječne mjere: visina 162—165,6 cm, indeks glave 76,3 do 78,4. Ova rasa spada južnom pojasu Evropida, koji počinje na istoku s Indidima, pa se nastavlja u Orijentalide i dalje u Mediteranide, jedan danas izolirani odvojak bili bi Polinezidi. Cijeli pojas ima kao zajednička obilježja dolihocefaliju i tamne kompleksije. Napose postoji, što se tiče kostura, velika sličnost ili gotovo istovjetnost između Orijentalida i Mediteranida, samo mekani dijelovi pokazuju bitne razlike.
Niži, ali vitak i gracilan stas, vrlo harmoničan u svojim proporcijama, koje gotovo točno odgovaraju umjetničkom kanonu, odlikuje sve te tipove. Usko, fino modelirano lice, nos ravan, srednje veličine, mala usta s punim usnama, jako naglašen bradni šiljak, lične kosti u istoj ravnini s uglom donje čeljusti, tako da uopće ne iskaču, obilježuju fiziognomiju Mediteranida. Sve je harmonično, ništa nije nezgrapno. Put je svijetlo smeđa s ružičastim tonom; na suncu jako potamni, tako da nije mnogo svjetlija od indidne. Kosa je valovita, tamno smeđa, a može biti gotovo crna s modrikastim ljeskom.
Glavno su područje Mediteranida obale Sredozemnog mora, Egipat, Libija, Tunis, Alžir, Maroko, Španjolska, južna Francuska, južna Italija i Grčka; jače je taj tip zastupan i u zapadnoj Engleskoj, a razasut je u većem ili manjem postotku kao rasna komponenta diljem cijele Evrope.
Već je Deniker razlikovao tri tipa: ibero-inzularni, atlanto-mediteranski i berberski, koje danas podređujemo pojmu mediteranidne rase. Haddon (1924) govori o starijem robustnijem eurafričkom sloju i mlađem sredozemnom. Svakako ima razlika između mediteranidnih tipova. Profinjeni tip nalazimo na sjevernom rubu Sredozemlja i odanle sve do Engleske. Na pojedinim područjima, u Liguriji, sjevernoj Španjolskoj, pa i ponekuda u Sjevernoj Africi ima uz to tip nižeg stasa i grubljih crta, a napokon diljem Sjeverne Afrike sve do srednje Sahare tip srednjeg stasa i tamnije, često žućkaste puti, s pomiješanim orijentalidnim i negroidnim elementima. Eickstedt naziva ta tri tipa ili tri podrase gracilnim Mediteranidima, Eurafrikanidima i Saharidima.
Već u ledeno doba doprli su kao aurignacski ili brnski tip, ljudi sličnih, ali primitivnijih obilježja u Evropu. Na te se forme nadovezuju Evrafrikanidi, dok su ostali sadašnji Mediteranidi potomci vala, koji je tek u kasnije doba preko Mezopotanije i Egipta zaposio Sredozemlje.
Dinaridi (tab. IV. sl. 4—7).
Prosječne mjere: visina tijela 167,0—178,3 cm, indeks glave 84,6—88,6. U donekle sličnom odnošaju, kao Mediteranidi prema Orijentalidima, stoje Dinaridi prema Anatolidima (Armenidima), tako da Škerlj govori o dinarmenskoj rasi.
Tijelo je Dinarida visoko i vitko, noge dugačke i trup kratak, dok je kod Anatolida obrnuto. Lice je usko, čelo visoko, nos velik, orlovski, dakle u tupom kutu ukrivljen, ne zaokruženo ukrivljen kao kod Anatolida. Nazolabijalni nabori jako se ističu i daju muškom licu odlučan muškarački izraz, što djeluje na žene drugih populacija vrlo privlačivo. Uho je usko i vrlo veliko. Bradni šiljak jako iskače, tako da je lice oštro profilirano. Usne su tanke. Koža je svijetlosmeđe ružičasta i na suncu potamni, ali ne tako jako kao kod Mediteranida. Kosa je glatka, smeđa, ili tamno smeđa, dlakavost je tijela prilična, napose pak pokazuju brada i brkovi jaki rast. Šarenica je oka, koje leži duboko, smeđa, ali često i siva. Pigmentacija, dakle, nije tako jaka kao kod Mediteranida. No najtipičnija je značajka Dinarida oblik lubanje, koja je straga sploštena, tako da se pričinja, kao da je glava okomito odsječena, te se kontura zatiljka i šije nalazi u istom pravcu. Drugovrsne brahicefalne lubanje, na pr. alpidne ili baltidne, straga su zaobljene.
Dinaridi su najgušće zastupani na zapadnom Balkanu, dakle u Hrvatskoj, Crnoj Gori i zapadnoj Srbiji. Odavle sežu nešto više miješani preko Alpa do Dunava i srednje Bavarske. U Tirolu je također gušći centar. Druga grana seže preko Moravske u Šlesku i preko Karpata u Ukrajinu. U alpskim dolinama, pa već i djelomično na Balkanu, postoji tip s dinaridnim formama, ali svijetlim kompleksijama, svakako krajevni tip, nastao miješanjem Dinarida i Nordida, a možda i Baltida. Lebzelter je označio taj tip noridnom (Noricum-Štajerska) rasom. Kod Slovenaca i u Hrvatskoj češći je dinaridni tip tamne kose, ali svjetlije puti i modrih očiju; taj tip Škerlj naziva savidnim (savskim). Dinarska obilježja nalazimo uz to razasuta po cijeloj Evropi, napose srednjoj. Jedan dinaridni eksklave nalazi se u Biskajskom zalivu, to je baskičko područje Francuske i Španjolske, ma da su ondje Dinaridi jako pomiješani s Mediteranidima. Drži se, da je baskički jezik zadnji ostatak onog jezika, kojim su prvobitno govorili Dinaridi.
Dinaridi su prodrli već rano u Evropu. Možda već u srednje kameno doba, a sigurno u novo kameno doba. Kultura zvonastih čaša, koja spada u to doba, popraćena je lubanjama tipično dinaridnog oblika, straga sploštenima, kao u slici prikazana lubanja, koja je nađena kod Nordlingena. Ova lubanja potječe, doduše, iz ranog brončanog doba, ali se nalazila na mjestu, gdje je bilo naselje iz neolitske kulture zvonastih čaša, što možemo razumjeti tako, da su Dinaridi skoro preuzeli brončanu kulturu. Kultura zvonastih čaša može se pratiti sve do Engleske. Ukoliko je ova kultura povezana samo s Dinaridima, to bi značilo, da su oni postojali već u ono staro doba diljem cijele srednje i zap. Evrope. Poteškoća je u tome, što bi prema pretpovijesnim nalazima kultura zvonastih čaša došla iz sjeverne Afrike preko Španjolske u Evropu, a Dinaridi su očito došli iz M. Azije na Balkan i onda dalje. Svakako nam lubanje pričaju, da su Dinaridi bar već kojih 6—8 tisuća godina naseljeni u Evropi.
Alpidi (tab. IV. sl. 9 i 10).
Prosječne mjere: visina 161,4—165,9 cm, indeks glave 84,8—86,5. Naziv Alpida potječe odatle, što su prije zvali taj tip Dissentis-tip, po nekoj švicarskoj populaciji, koja izrazito pokazuje tu formu. Lapouge je stoga upotrijebio naziv Homo alpinus; prihvatio ga je Ripley, pa se tako to ime općenito udomilo, ma da tog tipa ima više izvan Alpa nego u alpskim zemljama. Deniker je upotrijebio naziv »race occidentale« (zapadna rasa). Da je Ripley i Dinaride podredio tom tipu, proizlazi samo iz nedovoljnog poznavanja prilika, jer Alpidi imaju osim tamnih kompleksija baš suprotna obilježja od Dinarida.
Stas je nizak, zbijen i bucmast, tijelo naginje ugojenosti. Trup je razmjerno dugačak, a udovi kratki. Lice je nisko i široko, glava okrugla, nos malen s konkavnim hrptom, tako da se vide otvori nosa sprijeda, pogotovu kod žena. Kosa je glatka i nešto svjetlije smeđa od dinarske, šarenica je smeđa ili siva, koža je blijedo svijetlo smeđa s nešto crvenila.
Alpidi se nalaze najčešće zastupani u pojasu, koji se proteže preko srednje Francuske, zapadnih Alpa, jugozapadne Njemačke, Češke i Slovačke, a jedan ogranak nalazimo u sjevernoj Italiji. I u Madžarskoj ima dosta tog tipa, isto u zapadnoj Ukrajini. Inače je taj tip rjeđe razasut po čitavoj ostaloj Evropi. Poseban odvojak Alpida, koji su prije češće držali zasebnom rasom mongolidnih obilježja, nalazimo kod Laponaca na krajnjem sjeveru Skandinavije (lapidna podrasa). I E. Fischer shvaća Laponce kao rano od mongolidne loze odcijepljenu granu. Ali Laponci nemaju zapravo nikakvih mongolidnih obilježja, nego samo obilježja Alpida u nešto pojačanoj mjeri. Manji su po stasu (157,4 cm), lice im je šire i još okruglije, kosa tamnija, dok je šarenica smeđa, a često i siva. Eickstedt, komu se i Weinert priključuje, svrstava ispravno Laponce među Alpide, i to kao skupinu, koja je nadiranjem Nordida već u pretpovijesno doba odrinuta prema sjeveru (tab. IV. sl. 8.).
Alpidi su doprli u Evropu još ranije nego Dinaridi. U spilji Ofnet ima brahicefalnih lubanja alpidnih obilježja, dakle iz srednjeg kamenog doba, koje leži za nama bar 8—10.000 godina.
Nordidi (tab. IV. sl. 11—13).
Prosječne mjere: visina 172,4—173,5 cm, indeks glave 77,8 do 78,2. Ova je rasa prije nazivana teutonskom, germanskom ili indogermanskom, dok nije Deniker upotrijebio naziv nordijska. Stas je visok, vitak, s razmjerno kratkim trupom. Lice je usko duguljasto, čelo ravno visoko, s nadočnim lukom, otvor vjeđa stoji ravno ili se nešto malo spušta prema vani, oko leži, kao kod Dinarida, duboko. Usne su uske, bradni šiljak jasno iskače, lične kosti na obrazima nisu primjetljive, nos je velik, uzak i ravan, katkada lako orlovski ukrivljen. Dugačka se glava straga izbočuje iznad šije. Koža je bez ikakvog pigmenta, zato je ružičasta, na suncu ne potamni, nego jako pocrveni, kod jakog se sunčanja upali, oguli, ali poslije postaje opet bijela. Kosa je bujna, zlatno plave boje, tako i brada te brkovi; kod djece je kosa bjelkasto plava. Šarenica je modra.
Kako već ime kaže, Nordidi se nalaze najviše u sjevernim dijelovima Evrope, u Englezkoj i Škotskoj, istočnoj Irskoj, Norveškoj, Švedskoj, Finskoj, baltijskim zemljama, sjevernoj Rusiji i na gornjem Donu, u Poljskoj, sjevernoj Njemačkoj, Holandiji i sjevernoj Francuskoj, a ima ih raštrkanih i pomiješanih diljem cijele Evrope do malih eksklava na Atlasu i na Kavkazu.
Ova je rasa bila prije proširenija, kako pokazuju lubanje iz povijesnih i pretpovijesnih vremena te različni stari likovni proizvodi. U svih se naroda, koji govore arijskim jezicima, ova rasa vidno ističe; stari Perzijanci prikazivani su kao plavokosi, isto stari Grci i Rimljani; za indijsku brahmansku kastu starih vremena poznato je, da je bila bjeloputna i plavokosa. Po tome se ispravno zaključuje, da su bijeli ljudi nordijske rase tvorci indoevropskog prajezika.
Nordijska je rasa doprla u Evropu valjda dosta kasno i zauzela sjeverne krajeve, odakle je u valovima navaljivala prema jugu; prije kojih 5000 godina probila je iz svog prvobitnog središta u srednjoj Aziji u Indiju. Ima i mnijenja, da se već rano razvila u sjevernoj Evropi pa da se odatle širila na sve strane.
Nordijska je rasa imala svoje prethodnike u Evropi već koncem ledenog doba u Cro-Magnonskim tipovima, koji su možda praoblik nordijskih. Neki, kao Weinert, zalažu se za to, da je nordijska rasa nastala uopće u Evropi iz Cro-Magnonske. Svakako, još danas ima ostataka Cro-Magnonske rase; to je dalidna ili falska rasa (tab. IV. sl. 14). Paudler (1924) je upozorio, da ima u Dalarnu u južnoj Skandinaviji poseban plavokosi tip, ali sa sivkastom šarenicom, tijela širih dimenzija i širokog uglatog lica. Sličan se tip nalazi češće i u Westfalskoj, a napose je zastupan na Kanarskim otocima među Guančima. Lubanja tog tipa podudara se s Cro-Magnonskom, te se očito radi o ostatcima tog evropskog prastanovništva. Tragova Cro-Magnonskog tipa nalazimo usto u sjevernoj Africi, u Alžiru i Tripolisu, što znači, da se proširila ta stara rasa iz svojih sjevernih obitavališta daleko prema jugu još na prekretnici diluvija.
Eickstedt smatra tu rasu granom nordidne te je zove dalo-nordijskom, dok Nordide evropskog sjevera i zapada svrstava u teuto-nordijsku podrasu, a Nordijce na sjeveroistoku, napose među Velikorusima i Mordvincima, u feno-nordijsku. Potonji imaju više uglate oblike, crvenkasto plavu kosu i vodeno modru šarenicu.
Baltidi (tab. IV. sl. 15 i 16).
Prosječne mjere: visina 165—166 cm, indeks glave 80,9 do 83,2. Već je Deniker, kao dobar poznavalac tipova na slavenskom istoku, razlikovao posebnu plavokosu i brahicefalnu »race orientate« (istočnu rasu), a R. Virchow (1874) i A. Retzius (1878) već mnogo ranije istaknuli su t. zv. tip iz Finske. Čepurkovski (1906) je taj tip nazvao valdajskim, a poslije su se Pöeh, Czekanowsky, Bunak, Nordenstreng i dr. složili u tom, da se radi o posebnoj rasi, te je ovo stajalište općenito prihvaćeno. Nordenstreng je upotrijebio naziv istočno-baltijska rasa, Eickstedt ju zove istočno-evropidnom, Bunak baltijskom, kojim se nazivom služimo i ovdje, jer je najjednostavniji i najjasniji. Neki su prije tumačili plavokose brahicefale kao križance Nordida i Alpida, ali kako je plava kosa recesivno svojstvo, iz takvog križanja ne bi mogli izaći u tako velikom postotku plavokosi potomci.
Tijelo Baltida pokazuje slične proporcije kao Alpidi; dugi trup i kratke udove. Glava je razmjerno velika. Čelo je široko, a i lice; donja je čeljust jaka i uglata. Budući da su jagodice pomaknute nešto naprijed, lice se pričinja plosnato. Nos je malen s tupim vrškom. Koža je bijela kao kod Nordida, samo ne prosijavaju toliko krvne žilice, pa koža ima žućkasti ton. Kosa je glatka, mekana, pepeljasto plava, a šarenica bjelkasto modra ili sivo modra.
Ova je rasa nazvana i »slavenskom«, jer se jedan dio njenog područja proteže preko slavenskih zemalja. Središte je te rase svakako Bijela Rusija i Poljesje. Proširena je čitavom srednjom Rusijom do Sibirije, a prema zapadu se proteže u Njemačku, ima je mnogo u Finskoj, Madžarskoj, u Hrvatskom Zagorju; u priličnom postotku nalazimo je uopće na čitavom području slavenskih jezika, a rjeđe raštrkanu diljem cijele Evrope kao rasnu komponentu svih evropskih naroda.
Prvi se tragovi baltidne rase javljaju već na prekretnici novog kamenog i brončanog doba. Ovamo spadaju lubanje tipa I iz Šleske i Češke, koje je opisao O. Reche, lubanja iz Langenloisa u Austriji, pa lubanje, koje je Czekanowsky opisao kao predslavenski tip. Istog su tipa lubanje iz novog kamenog i brončanog doba, nađene kod Kijeva. Svi ti nalazi nemaju još nikakve veze sa starim Slavenima, nego potječu očito od fino-ugrijskih plemena. I neke mlađe lubanje baltidne rase, kao one iz Malog Poloma kod Vjatke, koje potječu iz 9. st. poslije Kr., nađene su u finskim grobovima.
Baltidi i Stari Slaveni.
Baltidna je rasa, dakle, već pojedinačno dosta rano dopirala u Evropu, i to kao nosilac fino-ugrijskih etničkih elemenata. U većoj mjeri prodrla je u Evropu sa starim Slavenima. Najstarije lubanje, za koje se može donekle sigurno ustanoviti, da su pripadale starim Slavenima, potječu tek iz 10.—12. st. po Kr., jer su tek tada prešli stari Slaveni od spaljivanja na pokapanje mrtvaca. Dok u staro-germanskim grobovima nalazimo gotovo isključivo nordidne lubanje, u svim slavenskim grobovima pored nordidnih zastupane su i lubanje širokog lica, mesocefalne ili brahicefalne, koje pripadaju, kako je to napose istakla I. Schwidetzky, ponajviše baltidnoj rasi. Nema sumnje, da su i Praslaveni bili nordidne rase, ali što su njihovi grobovi mlađi, to više ima primiješanih baltidnih elemenata, a uz to ponekuda i drugih rasa. Jasno se to vidi iz poredbe prosječnog indeksa dužine i širine lubanja iz ranijih i kasnijih grobova, kako to pokazuje ova tabela:
| Etn. skupina |
starije nalazište |
indeks |
mlađe nalazište
|
indeks
|
| Česi |
Žalov |
73,8
|
Levý Hradec |
76,7
|
| Slovaci |
Uh. Skalice |
72.2
|
Dĕvín |
80,4
|
| Obotriti (Macklemburg) |
Alt Bartelsdorf |
75,4
|
Gamehl |
77,6
|
| Dregovići (Sjev. Poljska) |
Zemni kurgani |
74,6
|
kameni kurgani |
76,7 |
| Polani (Juž. Poljska) |
kurgani |
76,6
|
groblja |
77,6 |
| Severjani (Zap. Rusija) |
kurgani |
73,5
|
groblja |
80,1 |
Starija nalazišta s manjim indeksom (dakle dužim lubanjama) sadržavaju pretežno nordidne lubanje, napose to vrijedi za Uhersku Skalicu, gdje ima isključivo nordidnih lubanja. Iz te poredbe proizlazi, da su nordidni Praslaveni već rano sebi asimilirali finske etničke grupe baltidne rase. Ove su prvobitno zauzimale tek podređeni položaj, ali su vremenom sve više pronicale u gospodujući sloj i, valjda uslijed veće plodnosti, sve više zauzimale maha tako, da je njihov udio skoro iznosio do 50%.
Nešto nordidne krvi primili su Slaveni ponovno od nekih germanskih plemena. F. K. Wolff drži, da bi mogli doći u obzir germanski Bastarni, ali se čini, da su se ti sastali sa Slavenima samo nakratko i nato nastavili svojom seobom u južnu Ukrajinu, gdje tada nije bilo Slavena. Nešto su više možda pridonijeli Istočni Goti, kojima su bili neko vrijeme podložni Slaveni na gornjem Dnjepru. Ali i to gospodstvo nije trajalo dugo. Svakako su asimilirali zapadni Slaveni, kada su prodrli na zapad, ostatke ondješnjih germanskih plemena, što je bio možda još najjači ponovni priljev nordidne komponente. Napokon su nešto nordidnih elemenata pridonijeli Normani, koji su prodrli u Rusiju te joj dali ime (ruotsi »veslači«). Ali sav taj priljev germanskih Nordida nije mogao biti od većeg značenja. Kako ističe Schwidetzky-jeva, rasčlanjenost je starih Slavena s obzirom na udio Nordida usmjerena od istoka prema zapadu, dok se u pravcu, kojim su doprla među Slavene germanska plemena, ne opaža nikakvo pojačanje nordidne komponente. Po tom ona zaključuje, da nordidni temelj Slavena potječe od prvobitno nordidnih Praslavena te da je germanski priljev bio vrlo neznatan.
Pored Baltida stari su Slaveni primili skoro i druge rase, tako da se već rano među sobom razlikuju po rasnim komponentama. Alpidne primjese primili su u srednjoj Evropi; što je ima na sjeveru, radi se valjda više o Lapidima. Na jugoistoku i na istočnom Balkanu pristupaju mediteranidni elementi, dok je na zap. Balkanu onaj nordidno-baltidni sloj, koji je donio slavenski jezik, primio mnogo dinarske primjese. Kod lubanja iz Hrvatske iz 10. do 12. st. iznosi prosječni indeks još 78,8, a iz 13.—15. st. već 81,4.
V. Rasa i narod.
Nazivlje rasa načelno izbjegava upotrebu etničkih imena i lingvističkih oznaka za rase, jer je rasa sasvim drugovrsni pojam nego pojam naroda ili pojam jezične skupine. Narod je skupina ljudi sa zajedničkom tradicijom, zajedničkim kulturnim i drugim duhovnim dobrima, zajedničkim ekonomskim interesima, spremna da sačuva i dalje izgrađuje tu zajednicu, s težnjom za zajedničkom državnosti. Važno je zajedničko kulturno dobro jezik, ma da ima i slučajeva, da se pripadnici istog naroda služe različnim jezicima, a pripadnici različnih naroda istim jezikom, kao na pr. neki Grci u Anatoliji, kojima je materinji jezik bio turski, ali su ga pisali grčkim slovima, a glavno im je zajedničko dobro bila pripadnost grčkoj crkvi.
Rasa je pojam biološke klasifikacije, i gotovo nigdje na svijetu ne podudara se više područje neke rase s područjem nekog naroda, a evropski su narodi jedan konglomerat onih pet evropidnih rasa, samo što je udio pojedinih rasa kod svakog naroda drugačiji, kako to objašnjavaju sheme u donjoj slici, u kojoj su u svrhu zorne predodžbe prikazani prema približnoj procjeni udjeli pet glavnih rasa kod nekih naroda. Nema, dakle, ni romanske ni germanske ni slavenske rase, nego postoje samo jezici i narodi te oznake. Govoriti na pr. o romanskoj rasi isto tako je nesmisleno, kao da bi netko govorio o dolihocefalnim glagolima, radi se o disparatnim pojmovima. Svaki je evropski narod četverostrukog ili peterostrukog podrijetla, svaki ima rasnu komponentu, koja mu je donijela ime i jezik, ali i druge su komponente od jednake važnosti, katkada baš one odlučuju o duševnom stilu naroda.
Rasni sastav nekog naroda ima, naime, izvjesno značenje. Kaošto se rase razlikuju po svojim fizičkim obilježjima, naravno je, da se razlikuju i po duševnim kvalitetama; način duševne reaktivnosti različit je, ljudi različnih rasa reagirat će pod istim uvjetima raznoliko, što se očituje u njihovu različnom temperamentu i različnom duševnom stilu. Udio pojedinih rasa, koje surađuju kod sastava kojeg naroda, suodlučivat će, dakle, o kakvoći duhovnih dobara tog naroda i njihove izgradnje. Naravno, rasni je sastav naroda samo jedan od čimbenika uz niz drugih, ekonomskih, geografskih, socijalnih i dr.
Još jedan drugi biološki momenat proizlazi iz narodne zajednice. Narod je, naime, u izvjesnoj mjeri rasplodna zajednica, jer se u velikoj većini sklapaju brakovi između pripadnika istog naroda. Radi toga se sve više među sobom miješaju nasljedni faktori rasa, koje sačinjavaju narod, tako da postoji težnja, da se stvori neka specifička mješavina nasljednih obilježja. Nije tko samo unutar jednog naroda, nego se svaki narod sastoji iz više takvih rasplodnih zajednica. Svaki, donekle ograničen kraj takva je rasplodna zajednica, jer se većina brakova sklapa unutar toga kraja. Kako narod nije homogena mješavina rasa, koje ga izgrađuju, nego ima prema različnim krajevima različnog udjela pojedinih rasa, to će se kroz više generacija u svakom kraju razviti njima svojstvena mješavina rasa. Ali rasa je samo jedan od čimbenika, specifički uvjeti života u tom kraju također su vrlo važan čimbenik. Tako će nastati krajevni tipovi, kao što su na pr. Zagorci, Ličani, Dalmatinci. Svaka rasplodna zajednica s vremenom dovodi do takve tipizacije; ima i rasplodnih zajednica, koje nisu krajevno određene, nego pripadnošću jednoj vjerskoj zajednici, ili, što je u prijašnja vremena bilo vrlo važno, pripadnošću nekom staležu, napose plemićkom. Ako je rasplodna zajednica vrlo uska, mogu se javiti uz tipizaciju i degenerativne pojave.
Kakvo je biološko značenje takve rasplodne zajednice, najlakše se može razumjeti, ako uvažimo teoretski i istinski broj predšasnika svakog pojedinca. Teoretski bi morao svaki čovjek imati u svakoj ascendentnoj generaciji 2n predšasnika, ako n znači redni broj dotične generacije, dakle 2 roditelja, 4 djeda i bake, 8 pradjedova i prababa, 16 šukundjedova i šukunbaba, a prije 1000 godina, ako se na 100 godina računaju 3 generacije, dakle u 30. generaciji, morao bi svatko imati 230 = 1.073,741.824 predšasnika! To, naravno, nije moglo biti tako, jer u ono vrijeme svih ljudi na zemlji nije bilo toliko. U istinskom bismo rodoslovlju našli, da se isti predšasnik izvjesne generacije nalazi na više mjesta rodoslovlja, da postoji t. zv. gubitak predšasnika, a uz to da ima, što više idemo natrag, to više istih predšasnika u rodoslovljima različitih ljudi neke rasplodne zajednice. Teoretski bi moralo imati današnjih 6 milijuna Hrvata, a da nisu među sobom u krvnom srodstvu, prije 1000 godina astronomski broj od nekih 6 tisuća bilijuna predšasnika, a svih je Hrvata tada bilo jedva pol milijuna. Uz to je mnogo njihovih loza u toku godina izumrlo, tako da jedva kojih 100.000 Hrvata onoga doba dolaze u obzir kao predšasnici današnjih Hrvata, uz to možda isto toliko ljudi od susjednih naroda, a to znači, da su svi članovi jednog naroda među sobom u krvnom srodstvu, i to mnogo bližem nego s ostalim ljudima, a napose to vrijedi za krajevne ili staleške rasplodne zajednice. Tako su, dakle, u svakom narodu svi nasljedni faktori davnih predšasnika najintenzivnije ispremiješani, svaki član takve zajednice ima neke istovjetne faktore s drugim članovima.
Čimbenici, koji su već u davnini djelovali kod stvaranja rasa, stalno su i sada djelatni u svakom narodu i u svakom njegovom dijelu, ali civilizacija sprečava daljnji njihov utjecaj, tako da ima samo početnih pojava, krajevnih tipova, a da opet skoro nastane neka pregrupacija, pa tako se kao u kaleidoskopu stalno mijenjaju slike, koje se stvaraju od nasljedstva, okoline i prilagođivanja.
LIT.: G. Deniker, Les races et les peuples de la terre, Pariz 1926; T. Dobzhansky, Genetics and the Origin of Species, 1937 (njem. prijevod: Die genetischen Grundlagen der Artbildung, Jena 1939); E. v. Eickstedt, Rassenkunde und Rassengeschichte der Menschheit, Stuttgart 1934, 2. izd. 1937—42 (tek polovica općeg dijela); E. Fischer, Die Rehobother Bastards, Jena 1913; Isti, Spezielle Anthropologie und Rassenlehre. Kultur der Gegenwart, III. 5., Leipzig i Berlin 1923; Isti, Rassen und Rassenbildung. Handwörterbuch der Naturwissenschaften, VIII., Jena 1939; Isti, Die Erbanlagen der Rassen, Bauer-Fischer-Lenz, Menschliche Erblehre und Rassenhygiene, 4. izd. I., München 1936; K. Gerlach, Begabung und Stammesherkunft im deutschen Volke, München 1929; H. Günther, Rassenkunde Europas, treće izdanje, München 1929; A. C. Haddon, The races of man and their distribution, Cambridge 1924; Human Biology and Racial Welfare, ed. by E. V. Cowdry, New-York 1930; H. Lundborg, Rassenmischung beim Menschen, Bibliographia Genetica, VIII., Hag 1931; Isti, Rassenkunde des schwedischen Volkes, Jena 1928; G. Montandon, La Race, les races, Pariz 1933; S. Ratković, Što je narod, Zagreb 1935; W. Z. Ripley, The Races of Europe, London 1900; W. Scheidt, Allgemeine Rassenkunde, München 1925; J. Schottky, Rasse und Krankheit, München 1937; I. Schwidetzky, Rassenkunde der Altslawen, Stuttgart 1938; P. Steffan, Handbuch der Blutgruppenkunde, München 1932; B. Škerlj, Človek, Ljubljana 1932; F. Weidenreich, Rasse und Körperbau, Berlin 1927; H. Weinert, Entstehung der Menschenrassen, Stuttgart 1938; B. Zarnik, O rasnom sastavu evropskog pučanstva, Kolo MH, Zagreb 1927; Isti, Krv i rasa, Priroda XXX., Zagreb 1930.B. Z-k.
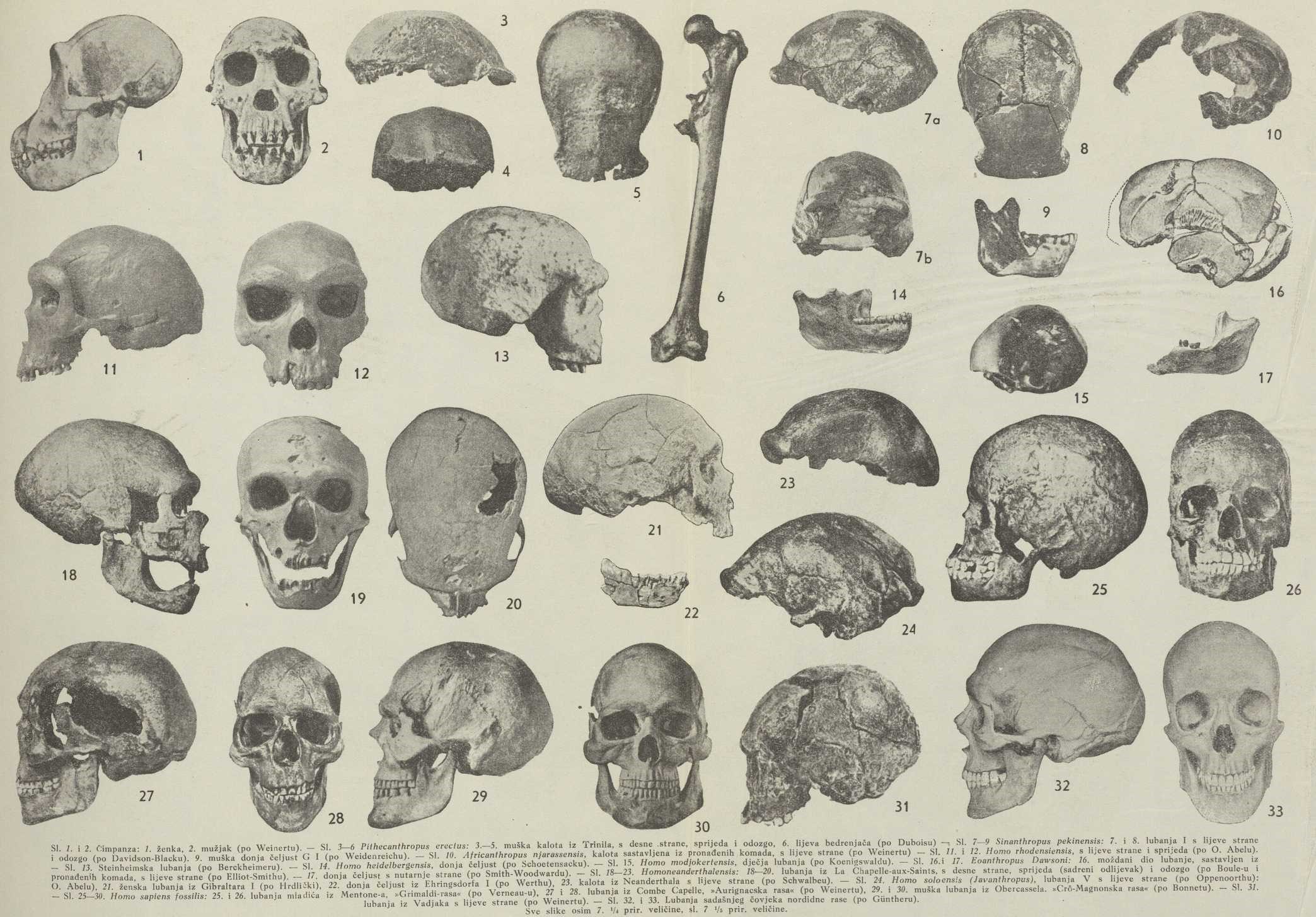 Prilog
PrilogČovjek u prehistoriji.
Vrijeme, kad se č. svojom kulturnom djelatnošću prvi put pojavljuje na zemlji, mogu približno ustanoviti paleontološka antropologija i prehistorija. Najstariji dosada nađeni ljudski kosturi i rukotvorine (artefakti) izravno upućuju na početak diluvija (pleistocena) antropozojskog razdoblja ili kvarternara (v.). Međutim, kakvagod se granica povuče između diluvija i prethodnog pliocena (v.), i makar se nijedan fosil čovjeka i njegovih rukotvorina izravno ne pridijeli pliocenu, ipak se opstojnost pliocenskog, dakle kenozojskog ili tercijarnog čovjeka, mora već sada smatrati sigurno utvrđenom. Oni, naime, tragovi čovjeka, koji u najmanju ruku pripadaju početnom, ili bar donjem diluviju, već su dosada veoma brojni i raštrkani na periferiji čitavog starog svijeta: u zapadnoj Evropi (Piltdown, Mauer-Heidelberg, Abbeville, Clacton-on-Sea), u istočnoj Africi (Kanam, Kanjera, Oldoway), na Javi (Modjokerto, Patjitan) i u Kini (Nihowan). Osim toga oni očituju znatne razlike u pogledu rase i tehničke djelatnosti. Zato su seobe iz zajedničkog središta (negdje u jugozapadnoj Aziji) do naznačene periferije te proces popratnih antropoloških i ergoloških diferencijacija zahtijevali barem toliko vremena, koliko je proteklo od srednjeg ili gornjeg pliocena do donjeg diluvija. To vrijedi i za slučaj, da se neki od navedenih nalaza iz donjeg diluvija djelomice osumnjiče u pogledu svoje geološko-paleontološke autentičnosti, jer ih još uvijek ostaje znatan broj jednako raštrkanih, koji su sigurno datirani. Jedinstveno pak podrijetlo ljudskog roda, barem u smislu monofiletizma, t. j. nazora o jednoj ishodnoj vrsti, usvajaju danas općenito i evolucionisti, a to neizravno pretpostavlja i neko ograničeno područje »prvih ljudi«, odakle su počele seobe njihovih potomaka (Weinert).
Priloženi pregled tipoloških i kronoloških rezultata paleontološke antropologije i prehistorije sastavljen je u okviru kvadriglacijarnog sistema (→ Kvarternar), tako da bi I., II. i III. interglacijar označivao jezgru ili središte donjeg, srednjeg i gornjeg diluvija ili pleistocena. U triglacijarnom sistemu, koji danas imade već mnogo pristaša, donji se pleistocen mora priključiti završnom ili uopće gornjem pliocenu.
A. JEDINSTVO PREHISTORIJSKOG I SADANJEG ČOVJEKA
Kad se promotre znatne razlike u izgledu fosilnih lubanja, moglo bi se činiti, kao da su to bitno različita bića. A ipak ih međusobno povezuje zajednička kulturna djelatnost. Ta se povezanost međutim očituje i u fizičkom pogledu.
Morfološki, t. j. po obliku, diluvijalne lubanje dadu se ponajprije razvrstati u dva glavna tipa: europoidni i pitekoidni. Europoidni je tip manje ili više sličan današnjim evropskim ljudima. Njegovi su stariji fosili iz donjeg i srednjeg diluvija nazvani prema pojedinim nalazištima (Ipswich, Piltdown i Swanscombe u Engleskoj, Kanam-Kanjera u istočnoj Africi, Capensis prema Cape Flats, uz druga mjesta, u južnoj Africi, Trinil na Javi), a mlađi fosili iz gornjeg diluvija zajedničkim imenom Homo s. cromagnonensis prema klasičnom nalazu u Cro-Magnon u pokrajini Dordogne u južnoj Francuskoj. Pitekoidni tip podsjeća u nekim crtama na majmune (Simiae) Anthropoidae ili Anthropomorphae (→ Antropoidni majmuni). U svom je izrazitom obliku nazvan Anthropus s podrazumijevanjem Pithecanthropus, t. j. »majmun-čovjek«, prema prvotnom nazivu glasovitog trinilskog fosila (E. Dubois 1892) i pobliže označen prema krajevima (Afrika, Kina ili lat. Sina, Java) i mjestima (Njarasa, Peking, Trinil-Sangiran-Kedung Brubus) pojedinih nalaza, a fosili uopćeno manje oštrih crta lubanje iz Evrope, Palestine (Galileje), Jave (Ngandong na rijeci Solo) i južne Afrike (Broken Hill u Rodeziji) nose zajednički naziv Homo neandertalensis ili naprosto Neandertalac, prema klasičnom nalazu u Neandertalu kraj Düsseldorfa u zapadnoj Njemačkoj. Ali, budući da se upotrebljavaju i posebni nazivi, H. galilaeensis za palestinskog, H. soloensis za javanskog i H. rhodesiensis za južnoafričkog Neandertalca, može se i evropski Neandertalac nazvati vlastitim imenom, i to H. krapinensis ili naprosto Krapinac, prema glasovitom nalazu u spilji na brijegu Hušnjakovu kraj Krapine (F. Koch i Gorjanović-Kramberger 1899—1905). Međutim sve brojniji fosilni materijal i njegovo točnije ispoređivanje već dovoljno opravdavaju, da se iz prvog i drugog tipa izluče neki nalazi kao poseban treći, australoidni ili posredni tip. To su uglavnom fosili ili neandertaloidni, t. j. više-manje slični Neandertalcu, ali toliko manje bridoviti u lubanji, da očituju također neke značajne crte europoidnog tipa, ili pak europoidni s toliko oštrijih crta, da se približuju Neandertalcu; zato se općeno podudaraju s fosilnim i današnjim Australcima (i s plemenima Ognjene zemlje), koji u sebi spajaju neandertalidne i europidne oznake (Klaatsch, Lebzelter, Škerlj, Weinert). Australoidni su fosili nazvani prema pojedinim nalazištima kao Modjokerto, Mauer-Heidelberg, Steinheim i t. d. (v. Pregled).
Stariji antropolozi, a i neki sadašnji (napose Weinert), stalno su nastojali, da u duhu strogog darvinizma morfološke tipove međusobno povezu u filetičke ili genealoške t. j. »razvojne stupnjeve«, koji bi se međusobno razlikovali kao vrste ili pače kao rodovi, i od kojih bi dva bila temeljna za smjer čitavog razvoja: Anthropus i Homo. Tako bi Javanthropus, čija je lubanja najbridovitija uopće, u smislu prvotnog naziva Pithecanthropus erectus bio pravi »missing link«, a čitava skupina Anthropus »stupanj prijelaznog oblika« između antropoida i hominida. Daljni bi »stupnjevi« imali biti: neki dosta neodređeni Homo praeneandertalensis kao Mauer-Heidelberg, Steinheim(!) i Broken Hill(?!), zatim »H. primigenius« t. j. Neandertalac prema staroj terminologiji, onda opet neki pobliže neodređeni H. postneandertalensis (djelomice bi ovamo pripadali australoidni fosili iz gornjeg diluvija), konačno »H. sapiens« (uglavnom australoidni i napose europoidni tip), i to najprije diluvialis (poglavito H. s. cromagnonensis) i za njim recens sa svim sadašnjim rasnim skupinama; između ovih bi australidi tvorili završnu točku fosilne »srednje linije« i polaznu točku takve recentne linije s izravnim nastavkom u europidima (bijelcima) i s »postranim linijama« u negridima (crncima) i mongolidima (žuta i bakrenasta rasna skupina). Tako Weinert. Međutim je jasno, da čitava konstrukcija mora pasti, ako pripadaju makar samo istoj geološkoj periodi oblici »stupnjeva« Anthropus i Homo, pogotovo »H. sapiens«. To će još više vrijediti, ako su ovi drugi od onih prvih ili »H. sapiens« od »H. primigenias«, odnosno australoidni Neandertalci od pitekoidnih, geološki još i stariji. A da je uistinu tako, očituje već letimična isporedba geološke kronologije fosilnih hominida u priloženom Pregledu. Zato se zadnjih godina sve više množe glasovi antropologa, koji ili u pojedinostima ili uopće naglašuju sinhronizam oprečnih tipova, odnosno anahronizam pretpostavljenih »razvojnih stupnjeva«. Evo ih nekoliko.
» ... Piltdown, kao i Kanam, pripada donjem diluviju. Srednjem diluviju pripadaju ... Ehringsdorf, Steinheim, Mauer-Heidelberg, Šukutien (Sinanthropus), Trinil (Javanthropus) i Kanjera, a gornjem diluviju ... La Chapelle-aux-Saints, La Quina ... (i ostali evropski Neandertalci) ... Tipovi Mauer-Heidelberg i Neandertalac (iz Ehringsdorfa i Steinheima), suvremenici Sinanthropusa i Javanthropusa, ne mogu biti njihovi potomci ...« (Hopwood prema L’Anthropologie 1936). — Javanthropus i Sinanthropus sigurno pripadaju samo srednjem diluviju i zato su suvremenici evropskih (starijih t. j. australoidnih) Neandertalaca, pa ne mogu biti njihovi pređi (van Stein-Callenfe1s 1936, Königswald 1937, Vaufrey 1938). — »Ne mogu, a da ponovno ne upozorim na nalaze Piltdown i Kenya (Kanam i Kanjera), koji ipak dokazuju u najmanju ruku istovremenost tipova Pithecanthropus i Homo ... (Odatle) se mora doći k logičkom zaključku, da Neandertalac ne predstavlja izravnog praoca, nego bratića naših pravih pređa. Mnogo je lakše priključiti Neandertalca skupini Pithecanthropus nego današnjeg čovjeka Neandertalcu« (Škerlj 1937). — »U Evropi je prije nekoliko godina lubanja, po Berckhemeru stručnjački iskopana kod Steinheima a. d. Murr, proizvela preokret u našim nazorima o razvoju diluvijalnog ljudskog roda. Ta lubanja, koja potječe iz početka trećeg glacijara (Riss), u mnogom je pogledu razvijenija nego Neandertalčeva, koju dosada poznajemo samo iz mlađih naslaga. S time je prviput sigurno utvrđeno, da su se u srednjem diluviju pojavili po vrsti ili rasi različiti hominidi. Mora se uočiti kao naročito sretna okolnost, da je ova spoznaja dobila daljnu potporu u jednom novom engleskom nalazu, u lubanji iz Swanscomba ... Prema gore spomenutim nalazima jedva se može sumnjati, da se razvoj čovječanstva nije odvijao po jednom stablu u ravnom pravcu, nego da su već zarana nastala cijepanja. Ako u srednjem diluviju dolaze s jedne strane oblici kao (javanski) Pithecanthropus i Sinanthropus, a s druge strane, samo nešto mlađi, takvi kao čovjek iz Swanscomba ili Steinheima, to se ne može držati, da su se ovi ljudski tipovi u tako kratko vrijeme razvili iz onih primitivnih oblika, naprotiv su ti različiti hominidi mogli biti suvremenici. Tome je u prilog također okolnost, da je pokraj Pithecanthropusa (na Javi) veoma vjerojatno već živio Homo, koji je, doduše, dosada poznat samo po svojim bedrenim kostima ... Dakle bi se stablo Homo već od ranog srednjeg diluvija samostalno dalje razvijalo pored stabla Anthropus. S tim se također ukazuje ponovno na obzorju Leakey-ev Homo kanamensis; njegov pronalazač svakako se čvrsto drži toga, da ostatci lubanja iz Kanama i Kanjere ... potječu iz netaknutih naslaga, dakle pripadaju najdonjem srednjem diluviju ili donjem diluviju ...« (Grahmann 1939). — »Što se tiče kronološke strane problema, čini se, da postoji nesavladiva poteškoća, ako se primitivni tipovi kao azijski prehominidi (Sinanthropus i Javanthropus) proglase »pređima« naprednijih hominida, kakve predstavljaju n. pr. ljudi iz Steinheima ili Swanscomba u Evropi, koji su živjeli otprilike u isto doba ...« (Weidenreich 1940). Istaknutu »nesavladivu poteškoću« ilustriraju Weidenreichove dvije popratne tabele. Na prvoj su tabeli hominidni fosili svrstani u »morfološkom slijedu«: prehominidi, Homo neandertalensis, H. sapiens intermedius (Steinheim i Swanscombe), H. sapiens fossilis (Piltdown, Kanam-Kanjera i dr.). Na drugoj su tabeli isti fosili poredani prema svom »kronološkom slijedu«: Piltdown i Kanam-Kanjera u prvom glacijaru Günz (odnosno u okviru triglacijarnog sistema na koncu pliocena!), Sinanthropus i Javanthropus u prvom interglacijaru Günz-Mindel i u drugom glacijaru Mindel (ali isp. prethodni tekst s onim, što ovdje slijedi, odnosno v. gore Hopwood, van Stein-Callnfels, Königswald, Vaufrey i Grahmann!), Swanscombe i Steinheim u drugom interglacijaru Mindel-Riss, najstariji pravi Neandertalci u trećem interglacijaru Riss-Würm ... (Weidenreich 1939 i 1940). Kronološki dakle slijed stavlja tako reći na glavu morfološki slijed, a lubanja iz Piltdowna s pravom nosi naziv »Eoanthropus« (čovjek u praskozorje), ali to je čovjek ne pitekoidnog nego europoidnog tipa (Weinert). — I sam je Weinert već g. 1938 morao s obzirom na H. modjokertensis iz donjeg diluvija priznati: »... Upravo je Java u zadnje vrijeme iznijela na svijetlo toliko novih fosila pračovjeka, da se time iznova zatalasala znanstvena rasprava o (Javi kao) klasičnom pretpovijesnom području. Ali uspjeh ovih novih plodova iskapanja na žalost je obratan od onoga, što bi se moralo očekivati... Nova su otkrića na Javi izazvala razilaženja u mišljenju, koja treba uzeti ozbiljno ... Moramo u najmanju ruku nabaciti i ostaviti otvorenim pitanje, da li su u diluvijalnim slojevima postojali ljudski oblici, koji dopuštaju tipove Anthropus, Neandertalensis i Sapiens diluvialis zamisliti i u drukčijem vremenskom slijedu«.
Međutim pored geološke kronologije, koja već sama za sebe stvara »nesavladivu poteškoću« u pretvaranju poredbenog slijeda morfoloških tipova u konstruirani slijed »razvojnih stupnjeva« i s obzirom na niže biološke tipove sili antropologe na usvajanje strmoglavih mutacija i nekih misterioznih »razvojnih težnja« (ortogeneze) uz sve veće ustupke konvergenciji t. j. genetički neovisnom razvoju sličnih oblika (tu se vidi presudna važnost geološke kronologije!), dakle pored toga treba još uočiti križanje varijacijskih raspona različitih tipova i oblika, i to naročito unutar pitekoidnog tipa te između njega i australoidno-europoidnog tipa. Odatle naime slijedi, da između njih ne postoji oštra granica, koja bi omogućila neko stupnjevanje na uštrb jedinstva vrste ljudskog roda u prošlosti i sadašnjosti. Sam Weinert, raspravljajući o pretpostavljenom »postneandertalskom stupnju«, mora ustanoviti: »... Kako na pojedinim lubanjama i kosturima tako i u cijelim skupinama nalazimo sad ovaj sad onaj dio kostiju još neandertalidan, dok su drugi (dijelovi) već u obliku od H. sapiens. Da se to ne pojavljuje istom u ovo doba (t. j. u gornjem diluviju), trebalo je uzgred spomenuti već kod nekih starijih nalaza; sjetimo se samo lubanje iz Steinheima. A isto tako se variranje između obadva stupnja (neandertalskog i australoidno-europoidnog) nastavlja dalje sve do današnjeg doba. Imade lubanja, koje bi se nesumnjivo uvrstile u neandertalski stupanj, kad njihove popratne okolnosti (t. j. geološko-paleontološki i prehistorijski podatci) ne bi jasno upućivali na kasnije doba. Zato sada ne možemo već neku pojedinačnu lubanju, koja očituje ovo ili ono neandertalsko obilježje, upotrijebiti kao provodni fosil... Ova se negativna konstatacija može doduše primiti kao neprijatna, ali ona se podudara s činjenicama, pa zato nema smisla, da se na tome štogod prevrće nasilno«. Dakle se varijacijski rasponi različitih tipova križaju i u gornjem diluviju i prije toga i poslije sve do danas. Zato Leakey (1936) zahtijeva »reviziju« neandertalskih fosila, jer da će se među njima naći više slučajeva oblika »Homo sapiens«, nego bi to neki htjeli priznati. A Cown i Keith (1939) su proučavanjem palestinskih »Neandertalaca« najprije izlučili dva oblika, nazvana prema poglavitim nalazištima Tabunu i Šukulu: prvi mlađi s neandertalskim, a drugi stariji s cromagnonskim vezama; kasnije su ustanovili, da je to ipak bio jedinstven narod, koji međutim u pojedincima očituje »sasvim neočekivano veliki varijacijski raspon« između Tabuna i Šukula kao dviju krajnjih varijacija. Weidenreich (1940) konstatira sasvim općenito: »Što više fosilnog materijala izlazi na javu, očituje se to veća varijabilnost i odatle to veća nesigurnost o tome, što treba proglasiti primitivnim«. A još je prije toga (1939) morao naglasiti: »... Homo sapiens, t. j. sadašnji čovjek, već je postojao u donjem diluviju Evrope (Piltdown), istočne Afrike (Kanam-Kanjera) i Male Azije (Palestina) i dalje kroz sve glacijarne periode. Još više može biti čudno, da taj H. sapiens nije za vrijeme ovakve opsežne ere iskusio nikakvih promjena u svojim morfološkim osebinama, dok su suvremeni oblici Anfhropus i H. primigenius (Neandertalac) bili podvrgnuti neprestanim i značajnim preobražajima«. To potvrđuje i Weinert svojim konstatacijama o pojedinim predstavnicima pitekoidnog tipa. Nesamo unutar čitavog oblika Anfhropus nego i među pojedincima jednog istog Sinanfhropusa, koji je poznat u većem broju iz glasovite spilje Šukutien (oko 200 km jugozapadno od Pekinga), postoje velike varijacije i u izvanjskom izgledu lubanje i u endokranijalnom kapacitetu (v. dolje). Oštre se crte ističu najviše kod Javanthropusa iz Trinila i Sangirana (u međusobnom razmaku od 50 km u srednjem dijelu Jave, vjerojatno najmlađi član!), a najmanje kod Africanthropusa (jezero Njarasa u pokrajini Tanganyika, vjerojatno najstariji član!), koji zalazi u varijacijski raspon Neandertalca i dapače u onaj australoidno-europoidnog tipa, jer očituje neke sličnosti s lubanjom iz Steinheima, dok u spilji šukutien imade zastupnika ovih relativnih krajnosti (Sinanthropus I i II) i različitih posrednih oblika, a među ove se može ubrojiti također Javanthropus iz Kedung Brubusa i po svom čelu i po svojoj donjoj čeljusti. Sve to samo potvrđuje iskustvo stečeno već s palestinskim »Neandertalcem«: što je veći broj poznatih pojedinaca određenog tipa ili oblika, to je veći varijacijski raspon njihovih značajnih crta. Zato su Javanthropus iz Trinila i Sangirana te Sinanthropus I samo jedna varijacijska krajnost i ništa više. Još veće varijacije očituje Neandertalac. Tako H. rhodesiensis (svakako najmlađi član, jer imade caries, pokvarene zube, što je sasvim nepoznato kod sigurno datiranih diluvijalnih fosila!) i H. soloensis iz javanskog gornjeg diluvija predstavljaju pitekoidni tip u velikoj mjeri sve do njegove krajnje varijacije u Javanthropusu. Naprotiv palestinski H. galilaeensis (osim kostura iz gornjeg sloja spilje Tabun) i H. steinheimensis (Weinertov poglaviti »Preneandertalac«), koji pripadaju srednjem diluviju, duboko zalaze u varijacijski raspon australoidno-europoidnog tipa, tako da se još jedva dadu uopće ubrojiti među Neandertalce (isp. gore Grahmann, Weidenreich, Cown i Keith); zato su u Pregledu stavljeni u red australoidnog ili posrednog tipa. Pravi pak evropski Neandertalac ili Krapinac varira u pojedincima između ovih relativnih krajnosti. Najbridovitija je lubanja onoga iz La Chapelle-aux-Saints iz zadnjega glacijara, a da se ipak ne izjednačuje s rodezijskim ili javanskim Neandertalcem, dok se neki stariji iz zadnjeg interglacijara kao fosili iz Ehringsdorfa i okolice Rima znatno približuju australoidnom tipu, odnosno steinheimskom i palestinskom »Neandertalcu«. Treći izraziti član tog tipa, H. modjokertensis iz javanskog donjeg diluvija, općeno se kreće u varijacijskom rasponu europoidnog tipa s pojedinačnim neandertaloidnim crtama, i to u manjoj ili bar nikako ne u većoj mjeri nego fosilni Austlci i lubanja iz Wadjaka na Javi ili današnji Australci, plemena Ognjene zemlje i pojedinci drugdje u Americi (na pr. iz algonkinskog plemena Piegan), koje Weinert sve bez izuzetka i bez ograničenja svrstava među »H. sapiens«.
Konačno svi su fosilni tipovi zajedno s današnjim različitim rasama jednako obuhvaćeni međusobnim križanjem varijacijskih raspona u pogledu endokranijalnog kapaciteta, t. j. obujma prostora za mozak, dakle za najvažniji organ u tijelu. To dokazuju slijedeći podaci uz prethodne brojke za najsavršenije majmune, sve izraženo u cm3.
|
Antropoidi:
|
| diluv. Australopithecus I 450; |
| „ Paranthropus robustus 600; |
| čimpanza najviše 440 (ž.) i 470 (m.); |
| gorila ,, 555 (ž.) i 585 (m.). |
|
Hominidi- di1uvija1ni:
|
| Piltdown 1070 ili 1300 (ili 1500?) |
| Swanscombe 1325 |
| Modjokerto 1100 |
| Steinheim 1070; |
| Africanthropus 1071 ili 1099 |
| Sinanthropus 850, 918 (ml. m.), 1050, 1100, 1150 (st. ž.), 1200, 1260. . . |
| Javanthropus: Trinil 950 (m.) i Sangiran 750 (ž.); |
| H. neandertalensis (inače rasa srednjeg rasta): |
| Gibraltar 1100 |
| Tabun (Palestina) 1260 |
| Ngandong (Java) između 1200 i 1300 |
| Broken Hill (Rodezija) 1300 |
| La Quina 1368 |
| prosječno 1400 |
| Neandertal 1408 |
| La Chapelle-aux-Saints 1626; |
| H. s. cromagnonensis (katkad do 2 m visine): do 1600; |
|
recentni:
|
| Evropljani uopće: prosječno 1300 ili 1350 (ž.) i 1500 (m.); |
| alpska rasa: prosječno 1333 (ž.) i 1429 (m.); |
|
,, ,, u Tirolu: varijacije između 880 i 1950;
|
| od izvanevropskih rasa visoka rasta: |
| stari Egipćani: prosječno 1336; |
| afrički crnci (i Melanezijci Nove Kaledonije): od 100 izmjerenih |
| 11 (6) između 1100 i 1300, |
| 45 (49) „ 1300 i 1500, |
| 41 (45) „ 1500 i 1700, |
| 3 (0) iznad 1700; |
| Melanezijci Nove Britanije: prosječno 1186 (ž.) i 1370 (m.); |
| Australci: prosječno 1154 ili 1146.4 (ž.) i 1310 ili 1295.3 (m.); od izvanevropskih rasa srednjeg rasta: |
| Malaji: prosječno 1223 (ž.) i 1414 (m.); |
| ,, na Javi: prosječno 1278 (ž.) i 1437 (m.); |
| od pigmoida: |
| Bušmani: m. prosječno 1240, ž. najmanje 900; |
| Vedda na Ceylonu: najmanje 960 (ž.?), odnosno i 918 (ž.); |
| od pravih Pigmejaca: |
| Andamanci na istoimenom otočju pred Birmom: prosječno 1148 |
| (ž.) i 1281 (m.). |
Iz navedenih podataka ponajprije slijedi, da od diluvijalnih vremena sve do danas razlika u endokranijalnom kapacitetu između ž. i m. iznosi 150 do 200 cm3. Nadalje se vidi, da se i danas u Evropi mogu naći pojedinci s kapacitetom lubanje od 880 ili samo 833 cm3: »Možemo primijetiti, u endokranijalnom se kapacitetu Pithecanthropus (trinilski Javanthropus) nalazi pače u krugu varijacije modernog čovjeka. Hechst ... izvješćuje o nekom duševno normalnom čovjeku, čiji mozak važe samo 788 grama ... (To znači), njegov je endokranijalni kapacitet morao iznositi oko 833 cm3. To je dakako izuzetan slučaj, ali sigurno endokranijalni kapacitet od 1000 cm3 dobro pristaje u granice normalne varijacije za modernog čovjeka« (Le Gros Clark 1937). A 1000 cm3 uistinu je prosječan kapacitet lubanje oblika Anthropus, jer se kreće između 750 i 1260 cm3, odnosno pretežna većina njegovih članova i k tomu svi suvremeni ili još stariji članovi australoidnog i europoidnog tipa nadmašuju taj prosjek. Evropski pak Neandertalac sa svojim maksimumom od 1626 cm3 (i to baš onaj najbridovitije lubanje iz La Chapelle!) i sa svojim prosjekom od 1400 cm3 naprosto je »hipermoderan«: nadmašuje naime prvom brojkom svog europoidnog suvremenika H. s. cromagnonensis, a drugom brojkom nesamo svoje daleke potomke Australce nego i neke druge recentne rase, među njima i visoko civilizirane stare Egipćane. Prema tome kapacitet lubanje kao njezina najznačajnija oznaka očituje kroz sve (Weinertove) »stupnjeve« i unutar pojedinih »stupnjeva« takove varijacije, da se nikakva brojka ne da odrediti kao isključiva svojina nekog »stupnja«, i da pogotovo ne može biti ni spomena o nekom uzlaznom stupnjevanju tog kapaciteta od oblika Anthropus preko Neandertalca do »Homo sapiens recens«.
Dodavši ostalo križanje varijacijskih raspona različitih tipova i njihovu geološku kronologiju, prema kojoj ovi »stupnjevi« uopće nijesu susljedni nego su većinom paralelni (ili pače »naopako susljedni«!), onda antropološka načela (isp. Kohlbrugge, Lebzelter, Weinert i dr.) traže, da se mjesto »filetičkih stupnjeva« upotrebljavaju neutralni nazivi kao tipovi, oblici i varijacije, odnosno kao temeljni rasni ogranci, rasne skupine i rase (Lebzelter). Sve navedeno naime dokazuje, da fosilni hominidi — i to jednako oni prognati (s izbočenim čeljustima) ili s niskim čelom ili s više manje istaknutim nadočnim i zatiljnim nabubrinama (u njima se ni sam Weinert pravo ne snalazi: sad su mu slučajne »hiperostoze«, onda opet »prave« izbočine, zatim jači »obrvni lukovi«, a kojiput samo nevažni »zafrkači«, sve to pak kolebanje radi »nezgodne« geološke kronologije dotičnih fosila!) kao i oni ortognati ili s visokim čelom ili bez frontalnih i okcipitalnih izbočina — stoje na istom »stupnju« sa sadašnjim čovjekom, i da između različitih fosilnih tipova i oblika ne postoje bitno veće razlike nego između današnjih rasnih ogranaka ili skupina.
Prema antropologu Lebzelteru među sadašnjim rasama postoje i takve morfološke razlike, da bi se po kriterijima biološke sistematike moglo govoriti nesamo o različitim vrstama nego pače i o različitim rodovima (na pr. između afričkog Pigmejca i Evropejca), a ipak danas nitko više ne poriče jedinstvo vrste svih živih rasa. Dubois pak, proučavajući kod različitih bioloških tipova t. zv. realnu cefalizaciju (omjer između veličine mozga i cijele muskulaturne mase), utvrđuje, da je ona kod Australca i Evropejca jednaka, dok je kod Neandertalca, Japanca i Eskima, kako se čini, veća nego kod Evropejca, pa odatle zaključuje: »To opet dokazuje, da čovjek nije postepeno napredovao u ljudskom ustrojstvu, nego da je nastao kao gotov rod (as one finished genus), i da uistinu svi mnogoliki tipovi čovjeka, u sadašnjosti i prošlosti, pripadaju jednom istom stupnju (to one and the same stage) prirodne animalne organizacije«. Dakle ne mogu ni članovi oblika Anthropus biti manje ljudi nego pripadnici pojedinih oblika i tipova Homo, niti Neandertalci opet manje nego »H. sapiens«, diluvijalni ili današnji. Tako se isti Dubois, zaslužan za otkriće trinilskog Javanthropusa, ne ustručava reći za Sinanthropusa, da je »istovetne rase s H. soloensis (javanskim Neandertalcem) t. j. veoma primitivan H. sapiens«. A to konačno i naročito potvrđuje njihova zajednička i istovetna kulturna t. j. duševna djelatnost (v. dolje). Uistinu su svi fosilni tipovi kao i sve današnje rase jednako »H. sapiens«. O tome naime, da li neko biće jest ili nije »sapiens« i prema tome uistinu čovjek, uopće ne može odlučivati antropologija proučavanjem kostiju, jer to mogu uglaviti samo prehistorija i etnologija proučavanjem kulture. Stoga bi bilo već krajnje vrijeme, da taj naziv »H. sapiens« u smislu ograničenog dijela ljudskog roda ili pogotovu nekog »stupnja« antropologija napusti, ako hoće ostati na svom području rada. S druge strane opet psiholog Bühler i za njim antropolog Lebzelter ističu: »...Čovjek ne biva prilagođen pomoću selekcije ili drugih bioloških faktora nego sam sebe prilagođuje proničući uvjete života. Time čovjek zauzimlje apsolutno izuzetan položaj u (biološkom) sistemu, te se vrsta i rasa pod utjecajem razuma izgrađuju uz sasvim druge preduvjete nego u ostalom živom svijetu«.
B. KULTURNA DJELATNOST PREHISTORIJSKOG ČOVJEKA. Ponajprije treba naglasiti, da sve današnje rase moraju pripadati istoj vrsti nesamo poradi neograničene plodnosti međurasnih brakova nego naročito i radi iste kulturne t. j. duševne djelatnosti. Kraj svih varijacija u kulturnim tipovima današnjih rasa temeljni kulturni elementi svima su zajednički: svi imadu neko gospodarstvo i neku ergologiju (oruđe, oružje, stan i t. d.), neku socijalnu organizaciju i neki jezik, neku umjetnost, znanost i religiju. Pače i u mnogim užim oblicima ovih temeljnih kulturnih elemenata postoji dalekosežna istovjetnost. Sve to dokazuje svima zajedničku istovjetnu ljudsku narav. Fosilne pak rase također jasno očituju kulturnu djelatnost, koliko su se ostatci i tragovi takve djelatnosti uopće mogli sačuvati u zemlji. Zato su nađeni većinom samo ostatci i tragovi elemenata materijalne kulture, i to naročito: upotreba vatre, izradba i upotreba oruđa i oružja od kamena, kosti i drva, pače u oblicima, koji se mogu susresti i kod današnjih rasa. To je već dosada nađeno kod svih fosilnih tipova, pa dokazuje nesamo bitnu istovjetnost kulturne djelatnosti njihove i današnjih rasa nego i njihovu pravu ljudsku duševnu, pobliže intelektualnu sposobnost. I najneznatnije naime oruđe, a pogotovu upotreba vatre (divlje se životinje boje vatre!), otkriva i zasvjedočuje ljudski duh, koji djeluje svijesno i shvaća odnos između uzroka i učinka, tako da taj odnos znade i praktički primijeniti kao odnos između sredstva i cilja. To je temeljna misao i najpriprostijih i najgenijalnijih otkrića. Nijedna životinja nije svijesno i praktički preskočila taj Rubikon (to priznaje i materijalist Bogoraz-Tan s obzirom na pokuse sa čimpanzama na Tenerifi!), te zato i nije proizvela kulture. Spoznaja principa kauzaliteta pak pretpostavlja stvaranje općih pojmova, koji su također temelj govoru i brojenju, a primjena principa uzročnosti izgrađuje govor i potiče nesamo na traganje za pojedinim uzrocima različitih pojava u prirodi nego također i na pitanje zadnjeg uzroka, i tako dovodi do ideje Najvišeg Bića t. j. do temelja religije, koja se uistinu nalazi, kao i govor, kod svih današnjih rasa, i to u takvom stanju, da pretpostavlja silno kompliciranu i dugu prošlost.
U pogledu sposobnosti govora kod članova fosilnog pitekoidnog tipa, kojima većinom sasvim manjka prednja izbočina donje čeljusti (imadu naime t. zv. negativni mentum), važna je izjava Weidenreicha, kustosa svih nalaza iz spilje Šukutien: »... Ne vjerujem, da previše riskiram, kad pretpostavljam, da se Sinanthropus već znao očitovati pomoću riječi. Mnoge indikacije, što ih pruža njegov kulturni život, podržavaju tu misao«. To vrijedi onda još u većoj mjeri za Neandertalca, a osim toga Weidenreich neizravno upozorava na prave kriterije za rješenje postavljenog pitanja: bitan je naime preduvjet za sposobnost govora ne dakako kakva kost, odnosno ovakav ili onakav oblik čeljusti, koja je uostalom »najvarijabilnija kost« (Virchow i Weinert), nego razuman i slobodan duh, a taj se očituje u kulturnom djelovanju uopće; zato bitnu sposobnost govora posjeduje i njemak, pa može svoje misli i nakane izražavati pismom ili drugim znakovima. Sinanthropus je također ostavio jasan trag svojih umjetničkih sposobnosti: komad šljunka s crtežima, koji prikazuju zdjelice; uistinu je on izrađivao i prave zdjele i čaše iz lubanje jelena (Breuil). A na kamenoj ploči, koja leži na neandertalskom kosturu u La Ferassie u Francuskoj, mogu se razabrati bojadisane crte: slabe su doduše, ali »bez maštanja, nego s nekom dobrohotnošću« dadu se u njima prepoznati orisi nekakve životinje (Comte Bégouen). U spilji Šukutien, gdje je Sinanthropus stanovao dugo vrijeme i ostavio iza sebe veliko ognjište uz obilato oruđe i oružje od kamena i kosti, nađene su i njegove vlastite kosti, ali najvećim dijelom samo lubanje i donje čeljusti, i to veoma brojne. U tom se pogledu njegova spilja upravo napadno podudara s naseljem današnjih Pigmejaca Andamanaca. Ovi naime obavljaju dvostruko sahranjivanje mrtvaca (→ Pokop) uz posebne obrede, koji očituju čvrstoću obiteljskih veza i veliku ljubav prema pokojnicima, a svršavaju s time, da nakon potpunog raspada leša najbliži rođaci uzmu i trajno na sebi nose ili kod sebe drže pokojnikovu donju čeljust ili lubanju, rijetko pak koju drugu kost; slične takve obrede imadu također Pigmejci Aeta na Filipinima, Tasmanci i jugoistočni Australci, a sve su to narodi, koji su nosioci najstarijih primitivnih kultura i među današnjim rasama zastupaju njihove najstarije ogranke. Na slične običaje upućuju nalazi javanskog Neandertalca (11 lubanja i k tomu samo 2 goljenice). Naprotiv su palestinski i europski Neandertalac većinom bili sahranjeni u pravim grobovima (zato je od njih nađeno dosta potpunih kostura); priloženo kameno oruđe uz nešto cijelih ili polomljenih životinjskih kosti kao ostataka hrane očituje vjeru u prekogrobni život. Ovakvi suglasni nalazi Neandertalaca iz različitih geoloških perioda u Palestini (u Šukulu čitavo groblje!) i na suprotnim krajevima Evrope (na Krimu, pa u Belgiji i Francuskoj) dozvoljavaju zaključak, da Neandertalčeve razbijene i porazbacane kosti u spilji kod Krapine najvjerojatnije upućuju na dvije mogućnosti: ili su grobovi bili raskopani od kasnijih stanovnika spilje, kako je to utvrđeno za spilju Kiik-Koba na Krimu, ili je Krapinac svoje mrtvace spaljivao, kako su to prigodice prakticirali današnji neandertaloidi t. j. plemena Ognjene zemlje i Tasmanci, a i neki Pigmejci Negrili u srednjoj Africi. Konačno je u spilji Šukutien nađeno na jednome mjestu zajedno više lubanja majmuna makaka. To napadno sjeća na ostatke lovačkih primicijskih žrtava glave (i velikih nožnih kostiju, odnosno mozga i moždine) spiljskog medvjeda u inventaru iz zadnjeg interglacijara u spiljama »Alpiena«, poglavito u Wildenmannlislochu i Drachenlochu u Švicarskoj i u Petershöhle u Bavarskoj; iste žrtve još danas prinose različiti narodi u arktičkom području Eurazije i Sjev. Amerike (napose Samojedi i Eskimi, i to poglavito od medvjeda i soba), zatim Vedda na Ceylonu i Pigmejci Negrili na Gabunu u Africi (ovi zadnji također od malih majmuna!). S tim je u vezi značajno, da Weidenreich nalazi na donjoj čeljusti i na gornjim sjekutićima Sinanthropusa neke osebine, koje da su posebno značajne za današnje Eskime, odnosno uopće za »mongolsku rasu«, kojoj se pribrajaju i Samojedi; povrh toga Sinanthropusa veže s Alpienom i s današnjim arktičnim prinosiocima spomenutih žrtava izrazita koštana industrija.
S obzirom na raznovrsno oruđe i oružje fosilnih hominida važno je istaći znatnu razliku ponajprije u upotrebi kosti i kamena, a onda u obradbi kamenog materijala (većinom kremena). Po svemu izgleda, da su članovi australoidnog tipa poglavito izrađivali koštano oruđe i oružje (isp. Mauer-Heidelberg i Steinheim). Najviše takvih rukotvorina imade u nalazištima, gdje dosada još nijesu otkrivene ljudske kosti (Nihowan u sjev. Kini i spomenuti Alpien u Evropi). U spilji Šukutien i koštani i kameni artefakti zastupani su podjednako (i to je značajno s obzirom na australoidne varijacije među pojedincima Sinanthropusa!). Naprotiv su sigurno članovi pitekoidnog i europoidnog tipa izrađivali svoje oruđe i oružje poglavito od kamena, ali na različan način.
Abbé Breuil, danas neosporni prvak u istraživanju protolitika ili starijeg kamenog doba (v.), dovoljnom je sigurnošću utvrdio, da od početka diluvija, i to svakako počevši od prvog interglacijara (prema kvadriglacijarnom sistemu), kamena industrija tog doba očituje dva i po izboru neposrednog materijala i po tehnici izradbe sasvim različita tipa, koji se paralelno i neovisno razvijaju kroz čitav diluvij sve do zadnjeg glacijara, tako da su njihovi nosioci sigurno morali biti dvije različite kulturne zajednice s vlastitim izgrađenim predajama, dvije različite grupe naroda i konačno dvije različite rase. Ovo zadnje poglavito zato: takva dugotrajna međusobna neovisnost razvoja dvaju tipova rukotvorina pretpostavlja i dugotrajnu izoliranost njihovih nosilaca, a to je temeljni uvjet za rasno diferenciranje (Lebzelter). K tome još treba pridodati različitu klimu i hranu jednih i drugih, bar u Evropi: jedni se pojavljuju u suptropskoj šumskoj klimi za punog zamaha svakog interglacijara; drugi pak u početku slijede glacijare u njihovu pridolaženju i odstupanju, t. j. zapravo prelazne periode ili subarktičku stepnu klimu, a iza trećeg glacijara ostaju također preko čitavog zadnjeg interglacijara i kroz veći dio zadnjeg glacijara. Prvi su dakle šumski interglacijalci, a drugi opet stepni glacijalci.
Bliže oznake spomenutih dvaju tipova protolitskih rukotvorina konačno omogućuju rasno identificiranje njihovih nosilaca. Šumski naime interglacijalci poglavito obrađuju jezgru kamena, a stepni glacijalci ovu zanemaruju i svoju tehničku vještinu posvećuju samo iverju, koje doduše upotrebljuju i oni prvi, ali bez posebne obradbe, jednako kao ovi drugi opet jezgru. Zato se prva industrija zove: nukleogena ili jezgraška, a druga: asulogena ili iveraška. Nadalje se jezgraši i iveraši međusobno razlikuju i u tehnici izradbe svojih rukotvorina. Jezgraši okresuju jezgru kamena preko obje strane ili plohe (odatle Breuilov naziv »bifaces«) i preko svih rubova, koji na taj način postaju oštri. Samo donji odeblji i oširoki kraj ostaje neokvrcan, pa je očito bio ili prihvaćan rukom ili nasađen na držak od roga, kosti ili drveta, već prema veličini i svrsi tog oruđa. Njegov gornji uski kraj svestranim okvrcavanjem prelazi (napose u kasnijem razvoju) u tanak i oštar šiljak. To je onaj tipični ručni klin (»Faustkeil«) u obliku jajeta, srca ili badema, koji je očito bio neki opći instrumenat: za rezanje i struganje, za ubadanje i bušenje, a također upotrebljavan kao oštrica strijele ili koplja, sjekirice i možda motike. Iveraši trebaju za iste svrhe čitav niz oruđa prema prikladnosti pojedinog ivera, pa tako kod njih nastaju različiti instrumenti najrazličitijih oblika: nožići, strugala, svrdlašca i drugi slični šiljci. Pri tom je dotični iver okvrcavanjem zaoštren obično samo na jednom uglu ili manjem dijelu rubova, a strane (plohe) ivera ili uopće nisu okvrcane ili je okvrcana samo jedna.
Jezgraška tradicija obuhvata tri glavne faze (nazivi potječu od klasičnih nalazišta poglavito u Francuskoj): Abbevillien (Chelléen) I-II u prvom, Acheuléen I-V u drugom i Micoquien u trećem interglacijaru. Iveraška tradicija počinje kao Clactonien I na izmaku prvog interglacijara sa približavanjem drugog glacijara te se pri njegovom odstupanju kod nadolaženja drugog interglacijara nastavlja kao Clactonien II, koji se dalje razvija u dvije odjelite linije: Levalloisien i Tayacien-Moustérien. Levalloisien I-II počinje na izmaku drugog interglacijara sa približavanjem trećeg glacijara i onda se pri njegovu odstupanju nastavlja kao Levalloisien III, da kao Levalloisien IV-VII postepeno proživljuje treći interglacijar i veći dio četvrtog glacijara. Tayacien I nešto se prije nego Levalloisien izdvaja iz Clactoniena na prijelazu iz drugog interglacijara u treći glacijar i pri njegovom odstupanju ostaje još kao Tayacien II, a iz njega se onda u trećem interglacijaru razvija Moustérien I, koji kao Mousterien II proživljuje prvi dio četvrtog glacijara.
Sada se konačno dade poduzeti rasno identificiranje jezgraša i iveraša. Prvi je Moustérien odao svog tvorca, i to Neandertalca. Ta je povezanost u Evropi već odavna uglavljena i općenito poznata, a sada je potvrđena također u Palestini. Javanski pak Neandertalac u gornjem diluviju svakako imade neku priprostu iverašku kamenu industriju, ali njezina udruženost s dotjeranim i tipološki značajnim koštanim rukotvorinama upućuje i na industrije miolitika ili srednjeg kamenog doba (van Stein-Callenfels), i to usprkos onih veoma oštrih crta lubanje tog Neandertalca. Trinilski pak kameni artefakti, koji su sigurno produkt iveraške tradicije, najvjerojatnije potječu od samog Javanthropusa. Uistinu su i ostala dva dosada otkrivena pripadnika rase Anthropus sigurno proizvodili asulogene kamene rukotvorine, i to: Sinanthropus gotovo sve njezine evropske varijante (Clactonien, Tayacien i Moustérien I, ali i druge tipove u vezi s bogato razvijenom koštanom industrijom, v. gore), a Africanthropus napose Levalloisien. Mora se dakle zaključiti, da su rase Anthropus i Homo neandertalensis t. j. pripadnici pitekoidnog tipa fosilnih hominida izvorni nosioci iveraške tradicije protolitskih industrija i kao takvi onda u Evropi također stepni glacijalci. Tvorci jezgraške kamene industrije dugo su vremena krili svoje prste. Ali već su odavna antropolog Keith i prehistoričari Bayer, Breuil i Bouyssonie izrazili svoju slutnju, da se iza nje krije »H. sapiens« t. j. europoidni tip fosilnih hominida. Uistinu su postepeno počeli izlaziti na javu članovi tog tipa različite geološke starosti i više ili manje sigurno datirani, a svi od reda povezani s nukleogenom kamenom industrijom, i to sa fazama Abbevillien i Acheuleen. To su nalazi: Piltdown i Swanscombe u Engleskoj, Kanam-Kanjera u istočnoj Africi (pokrajina Kenya) i H. capensis (napose Florisbad) u južnoj Africi; dakle opetovani nalazi iz dvaju kontinenata. Naročito je najnoviji nalaz, Swanscombe, koji je i sigurno datiran (drugi interglacijar) i sigurno povezan s obilnim inventarom starije jezgraške tradicije (Acheuléen III), definitivno potvrdio geološku i industrijalnu autentičnost svih ostalih. Zato treba zaključiti, da je europoidni fosilni tip izvorni nosilac jezgraške tradicije protolitskih industrija i kao takav onda u Evropi također šumski interglacijalac. Zato se dakle njegove kosti pronalaze razmjerno veoma rijetko:
na otvorenom zraku i u vlažnoj zemlji tropske ili suptropske šume one su se morale raspasti češće nego kosti pitekoidnog stepnog glacijalca, koji je većinom stanovao i mrtvace sahranjivao u spiljama.
S nalazima Kanam-Kanjera u najužoj je vezi Oldoway u susjednoj pokrajini Tanganyika, gdje su sve starije faze nukleogene tehnike zastupane u susljednim slojevima donjeg i srednjeg diluvija. Osim toga je u Patjitanu na Javi otkriven u slojevima, koji vjerojatno pripadaju donjem diluviju, tipični Abbevillien II uz nešto Clactoniena, a to se najbolje podudara s pojavom tamošnjeg H. modjokertensis iz donjeg diluvija, koji u sebi udružuje europoidne i australoidne crte. Obje su tradicije zastupane i u diluvijalnoj Australiji (Fürer-Haimendorf).
Na koncu se dade povući ovaj važan zaključak: ako izradba i upotreba oruđa već kao takvog nedvojbeno dokazuje intelektualne sposobnosti njegova auktora, onda navedene upravo kapriciozne razlike u izboru neposrednog materijala od iste sirovine i za iste svrhe kao i u tehnici obradbe s obzirom na akcidentalne okolnosti (okvrcavanje obiju ploha ili samo jedne, odnosno nijedne, što zapravo znači veće ili manje poljepšavanje oruđa!) isto tako dokazuje slobodnu volju njihovih majstora. A ta je također bar suodgovorna za izgradnju one tvrdoglave predaje, uz koju su jedni i drugi ustrajali kroz čitav dugi diluvij.
Navedeni podatci iz diluvijalne geologije, antropologije i prehistorije o šumskim interglacijalcima europoidnog tipa i jezgraške tradicije te o stepnim glacijalcima pitekoidnog tipa i iveraške tradicije upravo napadno potvrđuju ono, što je Lebzelter (1932) ustvrdio na temelju općih bioloških i antropoloških opažanja. On ponajprije ističe, da je fizička inferiornost čovjeka prema drugim biološkim tipovima upravo notorna: čovječje tijelo nije nikako specijalizirano za borbu s prirodom (to naglasuje i spomenuti Bogoraz-Tan). Iz utvrđene pak fizičke slabosti čovjeka zaključuje, da su prvi ljudi mogli živjeti jedino u tropskim i suptropskim šumama, koje su se još u diluvijalno doba povezano protezale kroz čitavu južnu Aziju sve do zapadne Afrike. Tu su prvobitni ljudi za prvu nuždu, dok još naime nisu dovoljno pronikli umjetno užiganje vatre, niti stvorili jače oružje i oruđe, mogli naći: u blagom podneblju zaštitu svoje golotinje i u ogromnom drveću zaklon od grabežljivih zvijeri, a u šumskom voću, korijenju i sitnim životinjama dovoljnu hranu; ukratko, tada su mogli biti samo šumski frugivori. Uistinu bi na otvorenim stepama tropskog ili suptropskog i pogotovu umjerenog pojasa brzo postali plijenom studeni, grabežljivaca i gladi. U ove su krajeve mogli ljudi prodrijeti i u njima se održati istom nakon toga, što su se jače umnožili, potpuno zagospodarili vatrom i proizveli takvo oružje i oruđe, da su bili sposobni braniti se od grabežljivaca i loviti veliku divljač, naročito stepne i tundrene travoždere, te se onda braniti njihovim mesom i odijevati njihovom kožom t. j. postati stepni karnivori. Uzevši zatim u obzir jedinstvo ljudskog roda, što ga općeno prihvaćaju moderni antropolozi, Lebzelter utvrđuje čisto biološkom poredbenom metodom one biljege, što ih je morala imati čovječja prarasa ili Forma typica prvobitnih šumskih frugivora. Ta je naime sigurno bila manje diferencirana i specijalizirana nego pojedini rasni ogranci, koji su iz nje proizašli do danas, napose oni fosilnog europoidnog i pitekoidnog tipa, jer u početku jedinstvenog ljudskog roda nisu mogle postojati takve velike opreke. Konačno Lebzelter ustanovljuje, da je fosilni europoidni tip (napose Piltdown) toj neutralnoj i nediferenciranoj Forma typica bliži nego pitekoidni tip, naročito oblik Anthropus, koji se previše jednostrano specijalizirao. Ovaj se dakle tip morao kasnije diferencirati iz Forma typica, i to seobom dijela prvobitnih šumskih frugivora u stepno područje, gdje je on konačno uslijed dugotrajne izolacije u stepama i uslijed prilagođivanja karnivornom (mesojednom) načinu života stekao nove rasne osebine (isp. »Postanak rasa« u prethodnom članku). Ručni klin, to najizrazitije oruđe jezgraške tradicije, Lebzelter izričito povezuje s poslovima frugivornog načina života šumskih interglacijalaca fosilnog europoidnog tipa. A onda bi se i ono znatno diferencirano oruđe iveraške tradicije dalo povezati s potrebama kompliciranijeg karnivornog života stepnih glacijalaca pitekoidnog tipa. Svakako se Lebzelterova teorija povrh svega toga dade dobro uskladiti s geološkom kronologijom članova ovih oprečnih tipova, a osim toga bi u Pregledu postavljeni fosilni australoidni ili posredni tip i njegova koštana industrija mogli u ocrtanom procesu uistinu označivati prelaznu fazu, sudeći napose po kronologiji i nekim drugim okolnostima njegovih najstarijih članova iz Modjokerta i Mauer-Heidelberga. Ova teorija također najbolje tumači ustanovljeno križanje varijacijskih raspona različitih tipova i oblika.
Što se konačno tiče miolitika ili srednjeg kamenog doba (v.), kojega su nosioci isključivo članovi europoidnog i australoidnog tipa t. j. europoidni »H. s. cromagnonensis« i njegove neandertaloidne varijacije (ipak isp. industriju javanskog Neandertalca!), ovdje je dosta istaknuti neka novija otkrića u kronološkom i tipološkom pogledu. Donedavna se o miolitiku, poglavito prema nalazištima u Francuskoj (odatle uopće francuski nazivi gotovo svih prahistorijskih industrija i kultura), sudilo uglavnom ovako: njegove su tri starije kulture t. j. Aurignacien, Solutréen i Magdalénien (s Azilienom) shvaćene kao susljedne i kao postepeni razvoj Moustériena II, pa su u tom smislu rasporedane kroz drugi dio zadnjeg glacijara i početak t. zv. postglacijara. Međutim su počela nadolaziti iznenađenja. U istočnoj su Africi Moustérien II i Aurignacien suvremeni. U Krasu, po istočnim rubovima Alpa (spilja Vindija u Hrv. Zagorju i cijeli niz spilja u Kranjskoj i Štajerskoj) i dalje na sjever u Moravskoj (tri spilje), Šleskoj i Galiciji Aurignacien se pojavljuje u zadnjem interglacijaru, dakle istovremeno s Moustérienom I i starijim Krapincem t. j. starijim pravim evropskim Neandertalcem. U spilji Pekárna u Moravskoj slojevi počinju s nekim surovim Aurignacienom I bez Moustériena, a također tu uopće nema Solutréena, nego se na Aurignacien II odmah nadovezuje Magdalénien. Najveće je iznenađenje donio Solutréen. U Francuskoj naime sve tri spomenute kulture pored koštanih rukotvorina i rezbarija, koje su i u Francuskoj i u drugim krajevima najobilnije i najdotjeranije u Magdalénienu, sadržaju izrazitu kamenu industriju iveraške tradicije, ali redovito s okvrcavanjem obiju ploha prema jezgraškoj tehnici; po obliku i svrsi oruđa i oružja svaka od ovih triju kultura imade svoje specijalitete. Tako je za francuski Solutréen značajan napose veliki šiljak (koplja) u obliku lovorova lista, veoma brižno i fino okvrcan s obje strane. Naprotiv je u brojnim spiljama Madžarske otkriven (djelomice suvremen s Aurignacienom) cijeli niz razvojnih faza tog šiljka i drugih solutréenskih rukotvorina, što je onda izraženo nazivima: prvobitni, rani, klasični i završni Solutréen. U prvoj se fazi nalazi još i pravi ručni klin protolitske jezgraške industrije, a istom u trećoj fazi dosiže veliki šiljak uglavnom (ipak ne sasvim) »klasičan« oblik i izradbu francuskog Solutréena, da konačno zahiri u četvrtoj završnoj fazi. Nadalje se u Předmosti i Brnu pored završnog Aurignaciena nalazi »moravski Solutréen«, koji se otprilike podudara s madžarskom drugom fazom. Nedavno je pak Hülle otkrio u Ilsenhöhle bei Ranis u Thüringenu u Njemačkoj zajedno i prvobitni i potpuno klasični Solutréen u zadnjem interglacijaru, i to ispod sloja s Moustérien I, dok istom iznad tog sloja slijede Aurignacien i Magdalénien kao u spilji Pekárna u Moravskoj. To znači pravu revoluciju u nazorima o evropskom miolitiku (Zotz), ali i u nazorima o geološkoj kronologiji europoidnog cromagnonskog čovjeka.
Iz navedenih se podataka dade naime ponajprije zaključiti, da su tri starije miolitske kulture prodirale u Evropu gotovo istovremeno, i to već u zadnjem interglacijaru i pri njegovu prijelazu u zadnji glacijar. A vjerojatno su dolazile s triju različitih strana: Aurignacien (tipični zastupnik iveraške tradicije) s juga ili jugoistoka, prvobitni Solutréen (djelomični baštinik jezgraške tradicije) s jugoistoka ili istoka i Magdalénien (osobiti zastupnik koštane industrije) sa sjevera kao preteča dansko-baltičkih i sibirskih kultura Kunda i Maglemose u t. zv. mesolitiku za vrijeme t. zv. postglacijara. U to se završno diluvijalno doba nadaleko po Evropi proširuju još dvije mesolitske ili mlađe miolitske kulture: Tardenoisien (mikrolitik »geometrijskih oblika«) kao nastavak aurignacienskog ogranka u sjevernoj Africi i Španiji (Capsiena) i Proto-Campignien kao jača obnova protolitske jezgraške tradicije (prema Menghinu) i djelomice uvod u neolitik ili mlađe kameno doba (v.) u aluviju ili geološkoj sadašnjosti. Nadalje iz navedenih podataka slijedi, da je svakako bar istovremeno s moustérienskim Neandertalcem u Evropi već živio tipični europoidni »H. s. cromagnonensis«. Uistinu su dijelovi njegova kostura i lubanje nađeni u Fürst-Johanns-Höhle kod Lautscha u Moravskoj uz interglacijarni Aurignacien (Bayer). Istom tipu pripadaju također nosioci Aurignaciena, Magdaléniena i njegovog degeneriranog ogranka Aziliena iz zadnjeg glacijara u Francuskoj, Belgiji i Bavarskoj. Australoidni tip, t. j. cromagnonskog čovjeka s nekim neandertalskim crtama predstavljaju nosioci starijeg Aurignaciena u Combe Capelle u Francuskoj, starijeg Magdaléniena u Oberkasselu kod Bonna u Njemačkoj i vjerojatno završnog Magdaléniena u spilji Biscordine u južnoj Francuskoj, a isto tako nosioci završnog Aurignaciena i ranog Solutréena u Moravskoj. Među nova iznenađenja svakako spada i to, da je na Javi s tamošnjim Neandertalcem H. Soloensis pored Moustériena I nađena industrija kosti i roga, koja u Evropi pripada Azilienu i t. zv. mesolitiku (Königswald i van Stein-Callenfels), dakle zadnjim izdancima miolitika u postglacijaru (zato opet ne bi više moglo iznenaditi, ako bi se u Evropi ili pogotovo u Palestini otkrio kakav Neandertalac kao nosilac nekih miolitskih kultura!). Tim je povodom engleski časopis »Nature« (prema L’Anthropologie 1936) pisao o potrebi »preispitanja« geološke kronologije tog Neandertalca najbridovitije lubanje (to svakako traži i njegov rodezijski srodnik s pokvarenim zubima, v. gore) dodavši: kao u slučaju Swanscomba i Piltdowna treba zaključiti, da je općenito prihvaćena filogeneza čovjeka »nesiguran kronološki vodič«, i da je »čovječji razvoj bio mnogo više zamršen, nego što se dosada mislilo«. Zato bi se s obzirom na razvoj čovječjeg tijela moglo s Lunnom, Köppelom i drugima zaključno reći: »zagonetka nije riješena«. Ali to je uistinu najmanje, što se reći mora, jer su morfološkim poredbenim putem konstruirani »razvojni stupnjevi« već postali neodrživi radi protivne geološke kronologije i radi križanja varijacijskih raspona različitih tipova, i jer kulturna t. j. duševna djelatnost u svakom slučaju otvara već od prvih poznatih početaka ljudskog roda nepremostiv jaz između čovjeka, zvao se on Anthropus ili Homo, i životinje (→ Duša).
LIT.: W. Schmidt, Völkerkunde und Urgeschichte in gemeinsamer Arbeit an der Aufhellung ältester Menschheitsgeschichte (Mitteilungen der Naturforschenden Gesellschaft Bern aus dem Jahre 1941), Bern 1942; H. Weinert, Entstehung der Menschenrassen, Stuttgart 1938; V. Lebzelter, Rassengeschichte der Menschheit, Salzburg 1932; E. von Eickstedt, Rassenkunde, 2. izd., Stuttgart 1939 i sl.; Die Kultur der Gegenwart, III. Teil, 5. Abteilung: Anthropologie, Leipzig und Berlin 1923; R. Grahmann, Neue Hominidenfunde und ihre Altersstellung (Forschungen und Fortschritte, 15. Jahrg., Nr. 26), Berlin 1939; Report on the Swanscombe Skull (The Journal of the R. Anthropological Institute, Vol. 68), London 1938; F. Weidenreich, The Sinanthropus Population of Choukoutien (Bulletin of the Geological Society of China, Vol. XIV, No 4), Peiping 1935; R. Vaufrey, Le Sinanthrope et la chasse aux têtes (L’Anthropologie, Vol. 48), Pariz 1938; R. von Königswald, Die fossilen Menschen von Java (Nachrichten der Deutschen Gesellschaft für Natur- und Völkerkunde Ostasiens, No 42), Bandoeng (Java) 1937; W. E. Le Gros Clark, The status of Pithecanthropus (Man, Vol. 37), London 1937; E. Dubois, On the Fossil Human Skulls recently discovered in Java and Pithecanthropus Erectus (Man, Vol. 37); R. Köppel, Zur Urgeschichte Palästinas, Rim 1937; F. Berckhemer, Der Urmenschenschädel aus den zwischeneiszeitlichen Fluss-Schotfern von Steinheim an der Murr (Forschungen und Fortschritte, 12. Jahrg.), Berlin 1936; B. Škerlj, Pithecanthropus und Homo (Ztscht. f. Rassenkunde, sv. 6, 3), Stuttgart 1937; M. Blažić, Evolucija i postanak čovjeka, Zagreb, 1939. — Abbé H. Breuil, Industries à bifaces et industries à éclats du Paléolithique ancien (Préthistoire, Vol. I/2), Paris 1932; H. Obermaier, Der Mensch der Vorzeit, Berlin 1912; O. Menghin, Weltgeschichte der Steinzeit, Wien 1931; H. Breuil, Le gisement à Sinanthropus de Chou-Kou-Tien et ses vestiges de feu et d’industrie (Anthropos, Vol. 27), Mödling-Wien 1932; H. Breuil, L’état actuel de nos connaissances sur les industries paléolitniques de Choukoutien et de Nihowan (L’Anthropologie, Vol. 45), Pariz 1935; P. V. van Stein-Callenfels, Recent discoveries of Skulls and Pleistocene Stone Implements in Java (Man, Vol. 36), London 1936; G. A. Bontsch-Osmolowskij, Die vorgeschichtliche Vergangenheit der Krym (Der Weltkreis, 2. Jahrg., Heft 9/10), Berlin 1931; H. Reck, Oldoway die Schlucht des Urmenschen, Leipzig 1933; E. Bächler, Das Alpine Paläolithikum der Schweiz, 2 Bde, Basel 1940; J. Andree, Der eiszeitliche Mensch in Deutschland und seine Kulturen (mit Beiträgen von W. Hülle . .), Stuttgart 1939; K. Absolon-R. Czižek, Palaeolithický výzkum ieskyně Pekàrny na Moravě (Die palaeolithische Erforschung der Pekárna-Höhle in Mähren), Brno 1932; S. Brodar-J. Bayer, Die Potočka Zijalka (Praehistorica, Bd I), Wien 1928; S. Brodar, Das Paläolithikum in Jugoslavien (Quartär, Bd I), Berlin 1938; M. Mottl, Faunen, Flora und Kultur des ungarischen Solutréen (Quartär, Bd I); P. V. van Stein-Callenfels, L’industrie osseuse de Ngandong (L’Anthropologie Vol. 46), Pariz 1936; D. Gorjanović-Kramberger, Život i kultura diluvijalnog čovjeka iz Krapine u Hrvatskoj, Zagreb 1913; A. Gahs, Krapinski čovjek i njegovi suvremenici, Zagreb 1928; A. G., Kopf-, Schädel- und Langknochenopfer bei Renntiervölkern (P. W. Schmidt-Festschrift), Wien 1928.A. G-s.
Čovjek u etnologiji → Etnologija → Kultura.
Čovjek u lingvistici → Lingvistika.
Čovjek kao duševno i duhovno biće. Dok se antropologija u prirodoznanstvenom smislu bavi tjelesnim ustrojstvom normalnoga čovjeka pojedinca uopće (fizička a.), i značajnim osobinama i razlikama ljudskih grupa napose (rasna a.), pravi je predmet čovjekova zanimanja za njegovo vlastito bistvo duševna priroda njegova ili antropologija kao teorija unutrašnjega života ljudskoga. U tom smislu po Kantu ona se odnosi na činjenice pojedinačne svijesti obuhvaćajući opću i posebnu psihologiju kao nauku o ljudskoj duševnosti za razliku od psihologije životinja, na pojave skupne svijesti pod imenom kolektivne psihologije »psihičke etnologije« (po Steinthalu), »psihologije narodâ« (po Wundtu) »socijalne psihologije« i sl. i na konkretno očitovanje duševnosti u životu i običajima pojedinih naroda pod imenom etnologije kao izražaja »narodnoga duha« u jeziku, pjesmi i priči, mišljenjima i vjerovanjima, u ćudorednim i pravnim nazorima i ostvarenju njihovu u društvenom redu i ustanovama, što se jednom riječi naziva narodnom kulturom. Na dnu svega toga zanimanja stoji svijesnoj prirodi ljudskoj svojstveni filozofijski nagon, kojemu početci idu u nedoglednu prošlost, na osvit duha do najranijih mitskih pogleda, štono sežu na kraj svijeta i do stana božjega. Metafizika (v.) i religija (v.) najdalji su horizonti mudrosti, osnovane na težnji čovjeka, da spozna sebe. Krajnji problemi ljudskoga bića i žića sačinjavaju filozofijski završetak znanja o čovjeku (filozofijska a.).
Najopćenitiji problem antropologije kao teorije unutrašnjega života čovjekova specifični je značaj njegove duševnosti u vezi s njegovim položajem u svijetu. Oduvijek je bilo među ljudima uvjerenje, da se čovjek odvaja od ostalih živih bića i odlikuje iznad svih. To uvjerenje, nije uzmaknulo ni onda, kad je K. Linnée prvi čovjeka svrstao u razred sisara i u red primata; nisu ga oslabili ni znanstveni rezultati o jednakosti anatomske građe s višim životinjama (»antropoidima«) i srodnosti fiziologijske funkcije, jer sve to, odnoseći se na fizičku prirodu, još nije odlučno za unutrašnje bistvo, osim ako bi se prihvatilo materijalističko stajalište, da je duševnost proizvod ili svojstvo tvari i duševna konstitucija naprosto određena tjelesnim, napose moždanskim ustrojstvom. Ni teorija evolucije ne može oduzeti snagu onom uvjerenju, jer i ona stoji pred činjenicom, da čovjeka od ostalih živih bića dijeli razmak, što preko značenja višega razvojnoga stupnja upućuje na osebiti stvaralački princip, po kojem čovjek stoji sam u svijetu. Neosporno je pri tom, da je čovjek u vremenskom redu pojave bićâ posljednji stvor, ujedno i najviši oblik postojanja, ne možda po tom, što bi u svakom pogledu bio najsavršeniji — naprotiv, različne životinje imaju bolje izgrađene pojedine organe, jače i oštrije razvite osjetne sposobnosti, dok je nagon u svih određeniji i sigurniji, bez obzira na to da ih drži u stalnom (stacionarnom) načinu života — ali po cjelokupnom sastavu bića, kako organskoga ustrojstva tako i duševne opreme, čovjek je vrhunac zemaljskoga stvorenja.
Na to ukazuje izraz, da je čovjek svijet u malom (»mikrokozam«) razumijevajući pod tim, da je u njem skupljeno sve, što ima u velikom svijetu: on je sazdan od tvari i, koliko je do toga, podvrgnut fizičko-kemijskoj zakonitosti i zavisan od uvjeta i utjecaja izvanjske prirode; u njega je živa sila, koju Aristotel naziva vegetativnom dušom, kakvu imadu već biljke, i po njoj je vezan na opći biologijski red (rađa se, hrani, raste, rasplođuje se i umire); s životinjskim se svijetom slaže u nižoj duševnosti, koju sačinjava osjetno zamjećivanje, čuvstvovanje i nagonsko teženje. Dovde ide prirodna stvarnost i njezino skupljanje u čovjeku. No s tim nisu iscrpljene mogućnosti ljudske, štoviše, istom preko te granice proteže se djelokrug, kojim samo čovjek vlada. Ta se vlast sastoji u sposobnosti razumnoga mišljenja i umnoga htijenja i uživanja, što se jednom riječi označuje kao duhovna snaga. Na toj se snazi, kako je učio već Aristotel, osniva drukčija vrst duševnosti, koja čovjeka odlikuje nad sva ostala živa bića. Iako se prema shvaćanju suvremene psihologije životinja ove ne mogu smatrati automatima, kojih bi se djelovanje sastojalo jedino u refleksnim kretnjama i slijepim nagonima, kako je mislio Descartes, duševni život njihov ne ide dalje od osjećanja i predočivanja s priključenim čuvstvenim dojmovima i instinktivnim, odnosno navikom ustaljenim reakcijama. Izgrađujući se na mehanizmu predočaba (→ asocijacija) životinjska je svijest skup procesa zamjećivanja (»perceptio«), pamćenja i sjećanja, koji stoji pod neposrednim utjecajem upravo aktualnoga utiska ili poriva (impulza), kreće se u odnosu sadašnjosti, koji se proširuje samo reproduciranim momentima prijašnjih iskustava i ne prelazi granicu pasivne (uzročno-posljedičnom nuždom određene), nevoljne djelatnosti.
Na osnovi tako određenih mogućnosti znadu i životinje doći ne samo do velikoga stupnja okretnosti i vještine, nego postići i znatnu gipkost u prilagođenju na nove prilike, lakoću i brzinu učenja i stvaranja nizova predočaba u asocijativnom postupku, ponavljanjem i vježbanjem, mogu donekle pokazati čak i svrhovite reakcije, koje čine utisak više duševne radnje. U povodu toga govori se gdjekada o inteligenciji životinja. No ne valja pregledati, kako sve to ponašanje, koliko ga ima, nastaje jedino u saobraćaju s čovjekom, u okolnostima i prigodama, koje je on stvorio ili namjestio, i da se stoga nalazi samo u pojedinom slučaju, a nije pojava čitave vrsti, koja bi nastajala iz unutrašnje potrebe dotičnih životinja i vodila ih do viših očitovanja. Opravdano se stoga duhovne sposobnosti ne mogu pridavati životinjama. Po Wundtu se u najboljem slučaju nalaze samo kod najviših životinja, i to tek u pojedinačnim zametcima i tragovima. Duhovni je život prednost pridržana jedino čovjeku, i ostaje to, sve ako bi se činilo, da se životinjska svijest ne da svesti na samo osjećanje i predočivanje, te bi se u povodu toga uzelo, da u njoj ima radnjâ, koje odgovaraju ljudskom razumu (koje su »analogon rationis«, kako je mislio Leibniz, a slično i mnogi psiholozi sve do danas), jer bi to bilo ne samo operiranje s drukčijim kategorijama nego bi vodilo i k snalaženju, koje nipošto ne dostiže snagu i značaj čovjekove duševnosti, koji je značaj Aristotel izrazio riječima: u duši je um.
Zanos ljudskoga života preko osjetno-nagonskoga očitovanja do umnoga vladanja ima tipično, za čitav ljudski rod mjerodavno značenje. Nalazi se svuda, gdje čovjek živi i radi, ne izuzevši ni narode na najnižem stupnju kulture, jer i oni u svoj zaostalosti svojoj pokazuju makar kako primitivne pojave i tvorevine duha, koje životinjama svakako nedostaju. Razlika od životinjskoga stanja već je na tom stupnju ljudskoga postojanja tako velika i duboka, da se i sa stajališta teorije evolucije (v.) mora ispuniti silnim vremenskim razdobljem, koje bi opravdalo razvitak višega duševnoga života. Općenito obilježje uzrasta na visinu umnoga držanja jest oslobođenje svijesti od sile časa i određivanje djelatnosti iz unutrašnje sabranosti i ustavnosti, čime se po E. Cassireru pasivni svijet utisaka preobražava u obrazovan (kultiviran) svijet stvaralačkoga izražaja. Osebitost čovjekova ogleda se tako u analitičko-sintetičkoj funkciji mišljenja (u razbiranju i shvaćanju), što vodi do stvaranja pojmova, kojima se mnoštvo iskustvenih podataka skuplja pod općim zajedničkim vidom (universalis consideratio); shvaćanje omogućuje pregled zbivanja u svijetu, izrađivanje slika o stvarima i događajima sa širega gledišta, nezavisno od slučajna opažanja i časovita raspoloženja, vezanje smišljeno utvrđenih uviđaja po sadržaju i odnosima u nizove i sustave sadržaja znanja. Sposobnosti »stvarati ideje« kao nalazu teorijskoga, u razmatranju i spoznavanju zabavljenoga duha odgovara na praktičnoj strani, odnoseći se na čuvstveno i voljno držanje, sposobnost razvijati duševnu snagu u svijesnoj na mjeri i u zahtjevu valjanosti, mjesto određivanja pod pritiskom neposredna dojma, u provali ćudi i pustoj samovolji ocjenjivati mogućnosti i postavljati ciljeve djelovanja, ukratko, »upravljati se prema idejama i idealima«. Držanje u odnosu na vrednote i svrhovite izglede (teleologijski stav) vodi onda do sposobnosti »ostvarivati ideje« u umnim djelima i uredbama života. U svem tom stvaralačkom gledanju i upravljanju na vrednote i svrhe, kao i u djelotvornom držanju u odnosu na njih čovjek se otima prirodnoj nuždi, dovija se vlasti nad sobom, koja se očituje u procesu »uljuđivanja« (humaniziranja) kao i vlasti nad prirodom u podređivanju prirodnih sila ljudskim zahtjevima, prilagođenju prirodnih okolnosti potrebama i iskorišćivanju prilika na korist ljudskoga roda. Klasično je to prikazao Sofoklo u »Antigoni« u glasovitoj korskoj pjesmi, koja se počinje riječima: »Mnogo je silno, ali ništa nije silnije od čovjeka« (s. 332 i d.).
Uvjet duhovno uznesenoga i s pravom ponositoga življenja je ljudska svijest po ovim posebnim, samo čovjeku svojstvenim značajkama: 1. ona se nalazi kao »ja«, odnosi stanja i djelatnosti na sebe, zna ih kao svoje (»mojski« naglašene), razvija se do spoznaje sebe i do samosvijesna odlučivanja, izgrađuje se do izrazite ličnosti s određenim smislenim i vrednosnim tendencijama; 2. ljudska svijest je društveno određena, prema pojedinom »ja« stoje druga isto tako sebe svijesna bića, kojima kažemo »ti«, odnosno »vi«, i s kojima stojimo u duševnom saobraćaju, možemo se na osnovi naših doživljaja prenijeti ili zamisliti u njihovo stanje, s njima osjećati, biti drugari u životnoj muci i borbi, u povodu čega se prirodna (biologijska) zajednica proširuje u društvenu organizaciju, pojedinačna svijest obuhvaća osjećaje (»simpatije«) i obzire na druge i uključuje u sebi skupnu svijest; 3. ljudska je svijest historijski određena, nije samo individualno protkana prošlošću, već je i skupno pored organskoga pamćenja vrsti povezana tradicijom, oživljava se na uspomeni primjernoga rada i uspjeha prošlih generacija i, nadovezujući na nj, napreduje u iznalaženju i stvaralačkom obrazovanju mogućnosti opstanka sve dalje ponad prirodnoga toka života. Razvijanju živototvorne djelatnosti pod uvjetima svijesti koncentrirane u znaku »ja«, od silne je pomoći to, što se u osobnom dodiru ljudskih bića isprvično nagonski značaj izražajnih, napose glasovnih kretnja, najprije promijenio u sredstvo saopćenja doživljaja i pozivanja na saučešće, dakle u ulogu posrednika duševnih veza društvenih, a onda se povisio i do znaka, kojim se fiksiraju i predstavljaju misli, pojmovi i idejni zahtjevi i omogućuje sporazumno operiranje s njima. Dok je tako govor poslužio razvijanju ideja, koje je Aristotel kao misaono (logično) određivanje svijesti suprotstavio prirodnom (fizičkom) zbivanju, dotle je ostvarivanju ideja, kao tehničkom proširenju sposobnosti ljudske, poslužila ruka kao »oruđe svega oruđa« i upotreba vatre, koju je, prema simbolički značajnoj grčkoj priči, ljudima s neba donio Prometej. Umno i tehničko sviđanje glavne su poluge ljudskoga uzdizanja, upravo ustajanje na život, koji se odnosi na nešto, što kao ugledano ili poželjno treba da bude. U povodu toga život u vlasti ljudskoj nije tek nastavak ili dotjerivanje prirodnoga zbivanja, možda potencirani izražaj prirodne sile, nego je preobražaj djelatnosti u svrhovito određivanje i stvaranje novoga čovjeka, što duhom prožet živi prosvijetljenim i udostojenim životom. Otud se nadaje idealni značaj ljudskoga rada, koji se u odnosu na ideje opravdava smislom i vrijednošću, a ne pokreće se i ne određuje samo u nužnom slijedu učina iz uzroka. Udešavajući se u umnom vidu i zahtjevu ljudsko je stanje stvar revna i brižna nastojanja i, prema zalaganju, valjano i zaslužno djelo. Pojam čovjeka uključuje kultiviranje dostojanstva ljudskoga, koje je, s obzirom na raspoloživu građu (tjelesno-duševnu konstituciju i izvanjske prilike života), njegovanje rođenoga bivstva (cultura naturae), a s obzirom na cilj njegovanje je duha (cultura animi). Po razvijanju duhovnih značenja na prirodnoj podlozi istom postaje čovjek čovjekom, što se označuje izrazom »uljuđen čovjek, čovječan čovjek«. Pregalački značaj toga postojanja čini, da je ljudsko stanje uvijek u stvaranju, nije siguran i stalan posjed, nije mirno uživanje, ne može se imati nego se mora uvijek nanovo osvajati i svagda je na cilju toliko, koliko je umna zanosa i ponosa uloženo u postizavanje njegovo, uzevši u obzir, da je u vezi s dinamičkim značajem duhovnoga bitka i to, da sadržaj i način života u dostojnosti ljudskoj, osnovan budući na uviđavnosti, nije naprosto dat, nije zakonom prirode određen ni zajamčen, već se mora tražiti i s uvažavanjem provoditi. U tom leži čitava problematika unutrašnjega čovjeka u jednu ruku kao psihologijsko pitanje o bivstvu ljudskoga duha (napose o funkciji mišljenja i volje, o razvitku vrednosne svijesti i savjesti, o samosvijesti i samoodređenju i o duševnom principu uopće), u drugu ruku kao pitanje o svrsi i vrijednosti ljudskoga nastojanja, odnoseći se na pojedine grane, kako teorijskoga tako i praktičnoga duha, na značaj i granice znanja (č. u spoznajnoj ulozi), zahtjeve moralne i religijske svijesti (č. kao ćudoredno biće) na osnove društvene obveze (č. kao društveno biće) na smisao povijesnoga zbivanja i kulturnoga stvaranja (problem filozofije povijesti i kulture). U rješavanje tih i drugih problema, što se tako pletu oko ljudskoga bića i žića, odvajkada zasijeca sudbonosna veza duše s tijelom, koja se od antropologijskoga značenja proširuje do kozmičke zagonetke (do pitanja odnosa duha i tvari) i postaje pitanjem čovjekova položaja i zadatka u svijetu. Stav prema tomu pitanju očituje se onda i u čuvstveno naglašenim pogledima, koji se kreću u protivnom smjeru. Na jednu stranu odrazuje se u njima tugaljivo raspoloženje i žalbe na nesreću i nevolju ljudsku, na poteškoće života i nedostatke prirode ljudske, po riječima Salustijevim, što je »slaba i kratka vijeka i više upravljana slučajem nego vrlinom«. Na drugu stranu odmnijevaju u zanosnom hvalospjevu o veličini i vrijednosti čovjeka, što u sebi gaji iskru božanskoga duha i u pouzdanju u konačnu pobjedu ideje prevladava teškoće položaja i tragiku vječite borbe. Čovjekom biti znači biti borac za duhovne vrednote (»militia vita hominis«). Prava je ljudska postojbina u carstvu duha.A. B-a.
Čovjek u umjetnosti → Umjetnost, Čovjek.